OBTENCION Y ANALISIS DE UNA MUTANTE DOMINANTE NEGATIVA DE LYT1 DE TRYPANOSOMA CRUZI
(especial para SIIC © Derechos reservados)
Coautor
John Swindle*
Infectious Disease Research Institute, Seattle, Washington, EE.UU.*
Recepción del artículo: 23 de junio, 2003
Aprobación: 0 de , 0000
 Conclusión breve
El proceso de infección de Trypanosoma cruzi es complejo, y seguramente involucra la participación de muchas proteínas; sin embargo, en muy pocas se cuenta con las evidencias experimentales que lo confirmen.
Resumen
El proceso de infección así como el ciclo de vida de Trypanosoma cruzi se conoce desde hace varios años; sin embargo, algunos aspectos celulares y moleculares de tales procesos han sido elucidados recientemente, y otros aún están bajo controversia o sin estudiar. El proceso de infección de Trypanosoma cruzi es complejo, y seguramente involucra la participación de muchas proteínas; sin embargo, en muy pocas se cuenta con las evidencias experimentales que lo confirme. En el presente trabajo se realiza la caracterización inicial de parásitos que expresan una copia adicional de LYT1 cuya secuencia de nucleótidos se encuentra modificada, con un fenotipo igual al observado en parásitos knock-out. Los parásitos presentaron capacidad infectiva y lítica disminuida, mientras que su habilidad de transición de estadio mostró estar incrementada. Este fenotipo es consistente con el comportamiento de una mutante dominante negativa en el que el producto de la copia adicional de LYT1 alterada podría estar inhibiendo la función normal de la proteína LYT1 nativa o del complejo proteico al que pudiera estar asociada.
Palabras clave
Actividad lítica, infectividad, parásitos, diferenciación
Clasificación en siicsalud
Conclusión breve
El proceso de infección de Trypanosoma cruzi es complejo, y seguramente involucra la participación de muchas proteínas; sin embargo, en muy pocas se cuenta con las evidencias experimentales que lo confirmen.
Resumen
El proceso de infección así como el ciclo de vida de Trypanosoma cruzi se conoce desde hace varios años; sin embargo, algunos aspectos celulares y moleculares de tales procesos han sido elucidados recientemente, y otros aún están bajo controversia o sin estudiar. El proceso de infección de Trypanosoma cruzi es complejo, y seguramente involucra la participación de muchas proteínas; sin embargo, en muy pocas se cuenta con las evidencias experimentales que lo confirme. En el presente trabajo se realiza la caracterización inicial de parásitos que expresan una copia adicional de LYT1 cuya secuencia de nucleótidos se encuentra modificada, con un fenotipo igual al observado en parásitos knock-out. Los parásitos presentaron capacidad infectiva y lítica disminuida, mientras que su habilidad de transición de estadio mostró estar incrementada. Este fenotipo es consistente con el comportamiento de una mutante dominante negativa en el que el producto de la copia adicional de LYT1 alterada podría estar inhibiendo la función normal de la proteína LYT1 nativa o del complejo proteico al que pudiera estar asociada.
Palabras clave
Actividad lítica, infectividad, parásitos, diferenciación
Clasificación en siicsalud
Artículos originales> Expertos del Mundo>
página www.siicsalud.com/des/expertos.php/20148
Especialidades
 Principal: Infectología,
Principal: Infectología,
Relacionadas: Bioquímica, Medicina Interna,
Enviar correspondencia a:
Dra. Rebeca Manning-Cela. Departamento de Biomedicina Molecular. Centro de Investigación y de Estudios Avanzados del IPN. Av. Istituto Politécnico Nacional No. 2508 Col. San Pedro Zacatenco México, D.F. C.P 07360, Apartado 14-740 México
Isolation and Analysis of a Dominant Negative LYT1 Mutation in Trypanosoma cruzi
Abstract
The infection process as well as the life cycle of Trypanosoma cruzi has been studied for many years; however, many cellular and molecular events involved in these processes have only recently been described and others remain unstudied. The process of infection of Trypanosoma cruzi is complex and surely involves the participation of many proteins; nevertheless, in very little experimental evidence exists that outlines the role of any given protein. The present work characterizes parasites that express an additional mutated copy of LYT1 , copy gene required for efficient infection. The mutant parasites displayed a phenotype similar to that of parasites carrying a LYT1 deletion. These parasites exhibited diminished infectivity and hemolytic capacity whereas their ability to cycle through the different developmental stages was enhanced. This phenotype is consistent with the behavior of a dominant negative mutation in which the product of the additional copy of altered LYT1 inhibits the normal function of native LYT1 protein or the protein complex it associates with.
Key words
Actividad lítica, infectividad, parásitos, diferenciación
OBTENCION Y ANALISIS DE UNA MUTANTE DOMINANTE NEGATIVA DE LYT1 DE TRYPANOSOMA CRUZI
(especial para SIIC © Derechos reservados)
Artículo completo
Introducción
Trypanosoma cruzi es el agente etiológico de la enfermedad de Chagas o tripanosomiasis americana, la cual afecta a mas de 20 millones de personas en Sur y Centro América. Tiene un ciclo de vida bifásico que incluye cuatro estadios de desarrollo diferentes, dos en insectos hemípteros de la familia Reduviidae y dos en el hospedero mamífero. En el insecto, la forma flagelada del parásito llamado epimastigote prolifera en la parte media del intestino antes de diferenciarse en tripomastigote metacíclico (forma infectiva y no replicativa del parásito) en el recto del insecto. Después de introducirse en la sangre del huésped, el parásito infecta las células de éste, diferenciándose en amastigote e iniciando nuevamente su replicación. Finalmente el amastigote se desarrolla en tripomastigote procíclico o de forma sanguínea, el cual sale de la célula y puede iniciar un nuevo ciclo de infección al infectar células vecinas o ser tomado nuevamente por el insecto durante la ingesta sanguínea transformándose nuevamente en epimastigote en el intestino de éste y completando así su ciclo de vida.1,2La invasión de la célula huésped por T cruzi es un evento complejo que involucra diversas etapas. Se inicia con la unión del parásito a la célula huésped, seguido de su internación, la formación de la vacuola parasitófora y finalmente su liberación al citoplasma de la célula donde se multiplicará para continuar con el ciclo biológico.3,4Durante el proceso de infección del parásito así como para completar su ciclo de vida indudablemente participan muchas proteínas; sin embargo, en muy pocas existen las evidencias experimentales que lo demuestren. Ejemplo de esto es TC-TOX, un factor del parásito que presenta actividad hemolítica en condiciones ácidas sugiriéndose su participación en el proceso de infección; sin embargo, esta evidencia es sólo indirecta, ya que está limitada a la demostración in vitro de su papel en la lisis celular permaneciendo aún oscuro si es o no requerida durante el proceso de infección.5,6 De igual manera se ha reportado una cis- o trans-peptidil-propil isomerasa secretada por tripomastigotes, de la que también se cuenta sólo con evidencias indirectas que sugieren su participación en la invasión de la célula huésped.7 Por otro lado, utilizando estrategias genéticas se demostró que la oligopeptidasa B de T cruzi tiene un papel importante en la invasión celular.8,9 Por último, en un trabajo previo mostramos que LYT1 es la segunda proteína que, sobre la base de evidencias genéticas, muestra ser requerida no sólo para una infección eficiente sino también en el proceso de transición de estadio del parásito. Usando metodologías genéticas se mostró que LYT1 no es esencial en epimastigotes; sin embargo, parásitos deficientes en LYT1 mostraron tres fenotipos sobresalientes. Estos parásitos son deficientes en infección, presentan desarrollo acelerado in vitro y actividad hemolítica disminuida en condicione ácidas.10 Análisis de RT-PCR sugirieron que fenotipos tan distantes posiblemente son el resultado de un trans-splicing alternativo regulado diferencialmente en los distintos estadios de desarrollo del parásito, lo cual da como producto tres ARNm distintos: dos de ellos codifican para la proteína LYT1 completa, y contienen una posible secuencia señal (SS) en su amino terminal, y un tercero codifica para la proteína incompleta carente de SS.11 Estos resultados son consistentes con la posibilidad de que sean producidas tanto una forma intracelular (citoplasmática o nuclear) como una forma secretada de la proteína, lo que podría explicar su participación en procesos tan diversos como la capacidad lítica en condiciones ácidas y regulación en la transición de estadio del parásito. El presente trabajo describe la obtención y análisis de parásitos que expresan además de los dos alelos silvestres de LYT1, una copia adicional modificada en su extremo 5\'. Los resultados mostraron que los parásitos presentaban disminución en su capacidad infectiva al igual que en su actividad hemolítica en condiciones ácidas, y capacidad de diferenciación de estadio aumentada. Este fenotipo sugiere que la mutación en LYT1 actúa como una mutante dominante negativa produciendo una proteína LYT1 mutada, que pudiera alterar el correcto funcionamiento del complejo proteico responsable de dichos fenotipos. Materiales y métodos
Células y parásitos. Fibroblastos 3T3 NIH se crecieron en medio esencial mínimo Dulbecco (DMEM) con 10% de suero fetal bovino, 1% de glutamina, y 5 µg/ml de penicilina-estreptomicina a 37 oC y 5% de CO2. Epimastigotes de T cruzi cepa Cl-Brener se cultivaron en medio de triptosa infusión de hígado (LIT) a 28 oC.12 Tanto tripomastigotes como amastigotes fueron obtenidos de sobre nadantes de fibroblastos 3T3 NIH infectados y fueron purificados utilizando un anticuerpo especifico contra el antígeno de superficie Ssp-4 de amastigotes (2C2B6).13 Obtención de la mutante dominante negativa. La construcción de los plásmidos pGEM5Zf(+)hyg::LYT1 y pGEM5Zf(+)neo::LYT1 (conteniendo la región codificante de hygr y neor y 714bp de la región flanqueante 5\' y 2,854bp de la región flanqueante 3\' de LYT1) así como los fragmentos digeridos con BglI utilizados para la electroporación y obtención de la mutante dominante negativa utilizada en este trabajo, son tal cual como se describieron anteriormente por nuestro grupo.10 Epimastigotes Cl-Brener fueron electroporados con 25 µg de cada fragmento purificado como se describió anteriormente12,14-16 y se seleccionaron después de 48 horas con G418 (250 µg/ml) e higromicina (250 µg/ml). Una vez obtenida la población resistente a los antibióticos, se obtuvieron clonas por dilución limitante en ausencia de la selección como se describió anteriormente.12Hibridación tipo Southern y Northern. Ambos métodos fueron realizados como se describió previamente.10,12 Las sondas moleculares utilizadas fueron obtenidas por PCR utilizando los iniciadores descritos anteriormente.10 Ensayos de infectividad. Monocapas de fibroblastos 3T3 NHI al 50% de confluencia fueron infectados con 2X106/ ml epimastigotes en fase logarítmica tal como se describió anteriormente.10 Debido a la ineficiencia en la metaciclogenesis mostrada por esta línea de parásitos iniciamos las infecciones con epimastigotes en lugar de tripomastigotes metacíclicos. El porcentaje de células infectadas se determinó bajo el microscopio, contando el número de células conteniendo parásitos con respecto al número total de células. Ensayos de transición de estadio. La transformación de epimastigotes a tripomastigotes metacíclicos o amastigotes fue determinado en ensayos in vitro tal como se describió anteriormente.10 El porcentaje de transformación se determinó bajo el microscopio contando el número de parásitos transformados con respecto al total.Ensayos de hemólisis. La actividad hemolítica de amastigotes y epimastigotes (2x107) fue evaluada en condiciones ácidas (NaOAc, pH 5.4 / 10mM; NaCl 200 mM; D-glucosa al 0.2%) utilizando eritrocitos de caballo (2x107) en un volumen final de 0.5 ml a 37 oC durante diferentes periodos de tiempo, como se describió anteriormente.10ResultadosObtención de la mutante dominante negativa de LYT1
Como se reportó anteriormente,10 con la finalidad de dilucidar el posible papel del producto del gen LYT1, en un trabajo previo se desarrollo una estrategia genética para la obtención de knock-outs de LYT1 de la cepa Cl-Brener de T cruzi utilizando recombinación homóloga sitio específica. Dicha estrategia dio lugar a clonas diferentes; una, en la que se llevo a cabo la eliminación de ambos alelos y otra en que la eliminación fue únicamente del alelo b resultando en la obtención de un knock-out doble (L16) y uno sencillo (L14) cuya caracterización inicial ya ha sido reportada.10 Finalmente se aisló una tercera clona denominada L13 que mostró tener un genotipo no esperado, en la que ambos alelos LYT1 fueron retenidos, pero además presentó un alelo LYT1 adicional. Como se muestra en la figura 1 el análisis tipo Southern mostró un patrón de restricción con Pst I que reveló las bandas de 3.7 kb, 2.5 kb y 1.2 kb correspondientes a los alelos a y b de LYT1 además de presentar una banda adicional de 1.8 kb que nos indicaba presentaba una secuencia adicional de LYT1 modificada en su extremo 5\'. Además, L13 retuvo el gen de resistencia a neomicina en una posición no determinada (dato no mostrado).
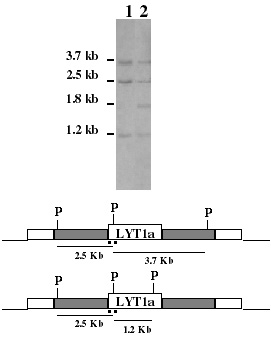
Figura 1. Análisis tipo Southern de la cepa silvestre Cl-Brener (carril 1) y la mutante dominante negativa L13 (carril 2). En la parte inferior del gel se muestra el patrón de restricción con PstI (P), que indica el tamaño de los fragmentos de restricción esperados para cada alelo y la ubicación de la sonda molecular utilizada (línea punteada).
El análisis tipo Northern, como se muestra en la figura 2 reveló que L13 expresa un ARNm de LYT1 de tamaño (1.8 kb) y niveles similares a los de la cepa silvestre. Además, dos bandas de menor tamaño y bajos niveles de expresión fueron observadas.

Figura 2. Análisis tipo Northern del transcrito de LYT1 de la cepa silvestre (carril 1) y la mutante dominante negativa L13 (carril 2). En cada carril se corrieron 10 µg de ARN total y se híbridaron con la sonda molecular de LYT1.
LYT1 mutada disminuye la eficiencia de infección in vitro
Ya que por nuestros resultados previos10 mostramos que LYT1 es importante en la infectividad del parásito; como primer punto, determinamos el comportamiento de la línea mutante L13 durante el proceso de infección de T cruzi. Como se muestra en la fgura 3 panel A, L13 mostró una cinética de infección deficiente comparada con los parásitos de la línea silvestre. En el panel B se muestra el promedio de 4 experimentos en el que se observa que en el momento en que la línea silvestre alcanzo el 100% de infección, L13 alcanzó solo un 5%.
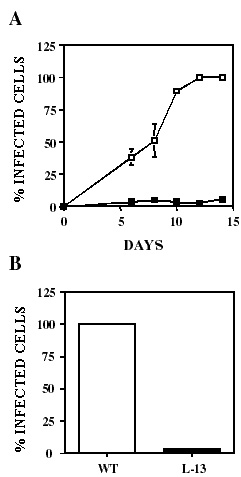
Figura 3. LYT1 mutada disminuye la eficiencia de infección in vitro. Fibroblastos 3T3 NIH se infectaron con parásitos silvestres (cuadros vacíos) o L13 (cuadros llenos) monitoreando el progreso de la infección a diferentes tiempos (A) así como en el punto máximo de infección (B). El porcentaje de células infectadas se determinó bajo el microscopio contando el número de células que contenían parásitos con respecto al número total de células. Los resultados son el promedio ± la desviación estándar de tres experimentos independientes.
Este resultado es inesperado ya que como se mostró en la fgura 1, L13 conserva los dos alelos de LYT1. Sin embargo, la presencia de una copia de LYT1 adicional que parece estar modificada en su secuencia en el extremo 5\' (fgura 1) daría lugar a un producto que podría estar compitiendo con las proteínas con las que interactúa LYT1 en la línea silvestre alterando de este modo su función y dando como resultado un comportamiento característico de una mutante dominante negativa. LYT1 mutada aumenta la eficiencia de transición de estadio in vitro
Para descartar la posibilidad de que la disminución en la eficiencia de infección fuera resultado de la incapacidad del parásito de transformarse para continuar con su ciclo de vida, evaluamos la habilidad de epimastigotes de L13 de transformarse in vitro a tripomastigotes o amastigotes. Como se muestra en la figura 4, la cinética de transición de estadio de L13 fue varias veces más eficiente tanto para amastigotes (panel A) como para tripomastigotes (panel B) comparado con la línea silvestre. En el primer caso alcanzó un 74% y en el segundo un 26% de eficiencia de conversión en su punto máximo (9 y 13 días respectivamente) comparado con aproximadamente un 3% en la línea silvestre.
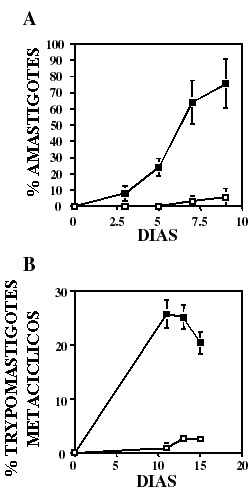
Figura 4. LYT1 mutada aumenta la eficiencia de transición de estadio in vitro. La eficiencia de transición de estadio de epimastigotes silvestres (cuadros vacíos) y L13 (cuadros llenos) a amastigotes extracelulares (A) o tripomastigotes metacíclicos (B) fue determinada en medio DMEM o LIT respectivamente. El porcentaje de parásitos en diferente estadio fue determinado bajo el microscopio contando el número de amastigotes o tripomastigotes metacíclicos con respecto al número total de parásitos. Los resultados son el promedio ± la desviación estándar de tres experimentos independientes.
LYT1 mutada disminuye la capacidad hemolítica del parásito en condiciones ácidas
A fin de evaluar si la disminución en la infectividad de L13 pudiera ser reflejo de una capacidad lítica disminuida, se llevaron a cabo ensayos de actividad hemolítica in vitro en condiciones ácidas de las líneas silvestre y L13 como se indicó en materiales y métodos. Como se muestra en la figura 5 hay una clara disminución en la capacidad hemolítica de amastigotes y epimastigotes de L13 comparando con la línea silvestre. El promedio de 3 experimentos indicó que en el punto máximo (3 horas) en que los amastigotes de la línea silvestre alcanzaron 81% de hemólisis, los amastigotes de L13 alcanzaron solo un 14% (panel A).
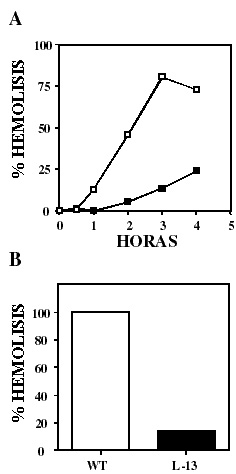
Figura 5. LYT1 mutada disminuye la capacidad hemolítica del parásito en condiciones ácidas. Se determino la capacidad lítica de amastigotes (A) y epimastigotes (B) de parásitos silvestres (cuadros vacíos) y L13 (cuadros llenos), evaluando el % de lisis de eritrocitos a 37 oC a pH 5.4 durante diferentes períodos de tiempo. El panel A es un experimento representativo de 3 independientes y el panel B es el promedio ± la desviación estándar de tres experimentos independientes. En cada punto se substrajo el valor de la lisis espontánea.
Si bien los amastigotes son el estadio que muestra mayor actividad hemolítica en condiciones ácidas, se ha observado ligera actividad en epimastigotes, por lo que también evaluamos este estadio. De igual manera, el promedio de 2 experimentos utilizando epimastigotes mostró que cuando la línea silvestre alcanza un 100% de hemólisis, L13 presenta solo el 14%.Discusión
Aún cuando el proceso de infección así como el ciclo de vida de este parásito se conocen desde hace varios años, diversos aspectos celulares y moleculares de dichos procesos han sido elucidados recientemente y otros aún permanecen bajo controversia o sin estudiar. Siendo el proceso de infección de T cruzi tan complejo, se espera la participación de diversas moléculas tanto del parásito como de la célula huésped. Sin embargo, a la fecha ha sido poco el avance en la dilucidación de las moléculas del parásito que participan en dicho proceso. Hasta ahora sólo han sido reportadas cuatro moléculas sobre las cuales las evidencias experimentales indirectas o directas las han presentado como involucradas en el proceso de infección del parásito. En el caso de TC-TOX5,6,17 y TcMIP7 se cuenta con evidencias indirectas, ya que en ambos casos está limitada a la demostración in vitro de su papel en la lisis celular o invasión celular respectivamente. En el caso de la oligopeptidasa B9 y LYT1,10 se demostró su participación en el proceso de infección del parásito utilizando estrategias genéticas. El presente trabajo es la continuación de dos trabajos previos, en el que reportamos la caracterización inicial de parásitos knock-out sencillos y dobles de LYT1. En estos informes se mostró que LYT1 no es esencial y que su ausencia está relacionada con atenuación en la capacidad infectiva de los parásitos, asociada con disminución en su actividad hemolítica en condiciones ácidas. La reintroducción de LYT1 restableció la infectividad de las mutantes nulas, confirmando el papel crítico que tiene LYT1 en la infección. También, experimentos de transición de estadio in vitro mostraron que los epimastigotes knock-out presentan capacidad aumentada de transformación a amastigotes y tripomastigotes metacíclicos en comparación con los parásitos silvestres, lo que sugiere que la infectividad disminuida observada no es el resultado de la incapacidad de los parásitos de completar su ciclo de vida.10 Estudios posteriores mostraron que la participación de LYT1 en fenotipos tan distantes podría ser resultado de un trans-splicing alternativo, el cual es regulado diferencialmente en los distintos estadios de desarrollo del parásito. Así, la presencia de los tres ARNm distintos encontrados es consistente con la posibilidad de que sean producidas tanto una forma intracelular (citoplasmática o nuclear) como una forma secretada de la proteína, lo que explicaría su participación en procesos tan diversos.11 Al igual que el doble knock-out de LYT1, L13, la clona en estudio en el presente trabajo, mostró los mismos tres fenotipos. Estos parásitos presentaron capacidad infectiva y hemolítica disminuida mientras que su capacidad de transición de estadio está incrementada. El genotipo de la línea de parásitos L13 fue inesperado, ya que en lugar de mostrar el reemplazo de LYT1 como se esperaría al presentar la resistencia a neomicina, presentó una copia adicional del gen. Sobre la base del análisis genómico, aparentemente esta copia adicional de LYT1 tiene una modificación en el extremo 5\' de su secuencia de nucleótidos; como se evidenció, al presentar una banda de hibridación de 1.8 Kb adicional a las de 3.7 Kb, 2.5 Kb y 1.2 Kb correspondientes a los alelos silvestres. Una posible explicación a estos resultados es que la copia de LYT1 alterada adicional se exprese y su producto actúe como una mutante dominante negativa inhibiendo la función normal de la proteína o del complejo proteico al que pudiera estar asociada. Por otro lado el análisis tipo Northern reveló que L13 expresa un transcripto de tamaño y niveles similares a los parásitos silvestres. Aún no sabemos por qué no observamos niveles alterados de transcrito de LYT1 en L13, aun cuando una copia extra de LYT1 se esté expresando. Las dos bandas de menor tamaño observadas podrían ser productos de degradación o transcriptos incompletos posiblemente resultado de la alteración en la secuencia de la copia extra de LYT1 encontrada en el análisis genómico; sin embargo, dilucidar esto requerirá mayor investigación. Los resultados mostraron que al igual que en los parásitos knock out la capacidad infectiva y hemolítica de L13 se vio comprometida. Nuevamente se observó que la cinética de hemólisis a pH ácido fue igual a la reportada para TC-TOX17 y knock-out de LYT1,10 proporcionando evidencias adicionales de que ambas moléculas están relacionadas al menos en la misma vía. También el hecho de que la habilidad de los parásitos L13 de diferenciarse in vitro está aumentada apoya la posibilidad de que en los epimastigotes LYT1 pudiera actuar como un supresor o regulador de diferenciación.En resumen, los tres fenotipos analizados mostraron comportamiento y niveles similares a los observados en la mutante nula; ello sugiere que dicho comportamiento corresponde a una mutante dominante negativa, en donde de alguna manera el producto de la copia de LYT1 alterada adicional podría estar inhibiendo la función normal de la proteína nativa. El análisis de dicha mutante no sólo confirma los hallazgos previos en los que se demuestra la participación de LYT1 en el proceso de infección y diferenciación de estadio, sino que también proporciona evidencias que apoyan la posibilidad de que LYT1 podría formar parte de un complejo proteico que participa en dichos procesos. Estudios posteriores serán necesarios para demostrar lo anterior así como para caracterizar las proteínas con las que interactúa.
Bibliografía del artículo
- Brener, Z. 1973. Biology of Trypanosoma cruzi. Annu Rev Microbiol; 27:347-82
- Dvorak, J.A. 1975. New in vitro approach to quantitation of Trypanosoma cruzi vertebrate cell interactions. New aprproaches Am. Trypanosom. Res. Sci. Pub 318:109-120.
- Rodríguez, A., Samoff, E., Rioult, M.G., Chung, A. and Andrews, N.W. 1996. Host cell invasion by trypanosomes requires lysosomes and microtubule/kinesin-mediated transport. J. Cell Biol. 134: 349-362.
- Tardieux, I., Webster, P., Ravesloot, J., Boron, W., Lunn, J.A., Heuser, J.E. and Andrews, N.W. 1992. Lysosome recruitment and fusion are early events required for Trypanosome invasion of mammalian cells. Cell. 71: 1117-1130.
- Andrews, N. W. 1990. The acid-active hemolysin of Trypanosoma cruzi. Experimental Parasitology. 71:241-4.
- Andrews, N. W., Abrams, C. K., Slatin, S. L. and Griffiths, G. 1990. A T. cruzi-secreted protein immunologically related to the complement component C9: evidence for membrane pore-forming activity at low pH. Cell. 61:1277-87.
- Moro, A., Ruiz-Cabello, F., Fernandez-Cano, A., Stock, R.P. and Gonzalez, A. 1995. Secretion by Trypanosoma cruzi of a peptidyl-prolyl cis-trans isomerase involved in cell infection. EMBO J. 14:2483-90.
- Burleigh, B.A., Caler, E.V., Webster, P. and Andrews, N.W. 1997. A cytosolic serine endopeptidase from Trypanosoma cruzi is required for the generation of Ca2+-signaling in mammalian cells. J. Cell. Biol. 136:609-620.
- Caler, E. V., Vaena de Avalos, S., Haynes, P. A., Andrews, N. W. and Burleigh, B. A. 1998. Oligopeptidase B-dependent signaling mediates host cell invasion by Trypanosoma cruzi. Embo J 17:4975-86.
- Manning-Cela, R., Cortes, A., Gonzalez-Rey, E., Van Boris, W.C., Swindle, J. and Gonzalez, A. 2001. LYT1 protein is required for efficient in vitro infection by Trypanosoma cruzi. Infect Immun. 69:3916-23.
- Manning-Cela, R., Gonzalez, A. and Swindle, J. 2002. Alternative splicing of LYT1 transcripts in Trypanosoma cruzi. Infect Immun. 70:4726-8.
- Hariharan, S., Ajioka, J., and Swindle, J. 1993. Stable transformation of Trypanosoma cruzi: inactivation of the PUB12.5 polyubiquitin gene by targeted gene disruption. Mol Biochem Parasitol. 57:15-30.
- Andrews, N,W., Hong, K,S., Robbins, E.S. and Nussenzweig, V. 1987. Stage-specific surface antigens expressed during the morphogenesis of vertebrate forms of Trypanosoma cruzi. Exp Parasitol. 64:474-84.
- Chung, S. H., Gillespie, R. D. and Swindle, J. 1994. Analyzing expression of the calmodulin and ubiquitin-fusion genes of Trypanosoma cruzi using simultaneous, independent dual gene replacements. Mol Biochem Parasitol 63:95-107.
- Gillespie, R. D., Ajioka, J. and Swindle, J. 1993. Using simultaneous, tandem gene replacements to study expression of the multicopy ubiquitin-fusion (FUS) gene family of Trypanosoma cruzi. Mol Biochem Parasitol 60:281-92.
- La Flamme, A. C., Buckner, F., Swindle, J., Ajioka, J., Barret, L. and Van, V. W. C. 1994. Engineering cytokine secretion from Trypanosoma cruzi. Memorias Do Instituto Oswaldo Cruz. 89:650-1
- Andrews, N. A. and Whitlow, M. B. 1989. Secretion by Trypanosoma cruzi of a hemolysin at low pH. Molecular and Biochemical Parasitology 33:249-256.
©
Está
expresamente prohibida la redistribución y la redifusión de todo o parte de los
contenidos de la Sociedad Iberoamericana de Información Científica (SIIC) S.A. sin
previo y expreso consentimiento de SIIC

