EFECTO DEL COLESTEROL DE LA DIETA SOBRE LA BIOSÍNTESIS DE LOS ACIDOS GRASOS NO SATURADOS
(especial para SIIC © Derechos reservados)
Coautores
Omar Jorge Rimoldi (Doctor en Bioquímica)* María Susana González (Licenciada en Ciencias** Ana María Bernasconi (Técnica Química)*
Instituto de Investigaciones Bioquímicas de La Plata (INIBIOLP), CONICET-UNLP, Facultad de Ciencias Médicas, La Plata*
Instituto de Investigaciones Bioquímicas**
Recepción del artículo: 25 de septiembre, 2002
Aprobación: 0 de , 0000
 Conclusión breve
En seres humanos o animales que están en el borde de la carencia de ácidos grasos esenciales, el 1% de colesterol es suficiente para conducirlos a una carencia efectiva al disminuir aún más los niveles de los ácidos araquidónico y docosahexenoico n-3.
Resumen
El agregado de 1% de colesterol a una dieta pobre en ácidos grasos esenciales (AGE) aumenta en la rata especialmente la proporción de los ésteres de colesterol tanto en las membranas microsomales como en los lípidos hepáticos totales. El aumento es, sin embargo, progresivo sólo en el hígado total. El colesterol dietario provoca tanto en estos animales como en los alimentados con dietas ricas en AGE un aumento de la actividad de la D9 desaturasa hepática así como del ARNm de la SCD-1. Ello produce un aumento en los lípidos hepáticos de los ácidos palmitoleico y oleico sintetizados por esa enzima. Este colesterol provoca también con ambas dietas un descenso de la actividad de las D6 y D5 desaturasas hepáticas, pero no se observó una variación del ARNm de la D6 desaturasa. Ese descenso conduce a una caída del ácido araquidónico hepático total y microsomal, y también a una disminución del docosahexenoico n-3 en lípidos totales del hígado. Ello demuestra que en seres humanos o animales que están en el borde de la carencia de AGE, el 1% de colesterol es suficiente para conducirlos a una carencia efectiva al disminuir aún más los niveles de los ácidos araquidónico y docosahexenoico n-3.
Palabras clave
Colesterol, ésteres de colesterol, ácidos grasos esenciales, efecto de la carencia, D6, D5 y D9 de, ARNm
Clasificación en siicsalud
Conclusión breve
En seres humanos o animales que están en el borde de la carencia de ácidos grasos esenciales, el 1% de colesterol es suficiente para conducirlos a una carencia efectiva al disminuir aún más los niveles de los ácidos araquidónico y docosahexenoico n-3.
Resumen
El agregado de 1% de colesterol a una dieta pobre en ácidos grasos esenciales (AGE) aumenta en la rata especialmente la proporción de los ésteres de colesterol tanto en las membranas microsomales como en los lípidos hepáticos totales. El aumento es, sin embargo, progresivo sólo en el hígado total. El colesterol dietario provoca tanto en estos animales como en los alimentados con dietas ricas en AGE un aumento de la actividad de la D9 desaturasa hepática así como del ARNm de la SCD-1. Ello produce un aumento en los lípidos hepáticos de los ácidos palmitoleico y oleico sintetizados por esa enzima. Este colesterol provoca también con ambas dietas un descenso de la actividad de las D6 y D5 desaturasas hepáticas, pero no se observó una variación del ARNm de la D6 desaturasa. Ese descenso conduce a una caída del ácido araquidónico hepático total y microsomal, y también a una disminución del docosahexenoico n-3 en lípidos totales del hígado. Ello demuestra que en seres humanos o animales que están en el borde de la carencia de AGE, el 1% de colesterol es suficiente para conducirlos a una carencia efectiva al disminuir aún más los niveles de los ácidos araquidónico y docosahexenoico n-3.
Palabras clave
Colesterol, ésteres de colesterol, ácidos grasos esenciales, efecto de la carencia, D6, D5 y D9 de, ARNm
Clasificación en siicsalud
Artículos originales> Expertos del Mundo>
página www.siicsalud.com/des/expertos.php/20330
Especialidades
 Principal: Nutrición,
Principal: Nutrición,
Relacionadas: Medicina Interna,
Enviar correspondencia a:
Dr. Rodolfo Roberto Brenner. Instituto de Investigaciones Bioquímicas de La Plata (INIBIOLP), CONICET-UNLP, Facultad de Ciencias Médicas, calles 60 y 120, (1900) La Plata, Argentina.
Dietary Cholesterol Effect on the Unsaturated Fatty Acid Biosynthesis
Abstract
The addition of 1% cholesterol to a diet low in essential fatty acids (EFA) specially increases in the rat the proportion of cholesterol esters in both total liver and microsomal membrane lipids. Yet the increase is only progressive in total liver. Dietary cholesterol provokes in these animals, as well as in animals feeding diets high in EFA, an enhancement of liver D9 desaturase activity and the SCD-1mRNA. This produces an increase in hepatic lipids of palmitoleic and oleic acids synthesized by the enzyme. Dietary cholesterol provokes also in the of both diets, a decrease of hepatic D6 and D5 desaturase activities, but there is no alteration of D6 desaturase mRNA. This decrease produces a fall of arachidonic acid in liver total lipids and microsomes, and also of docosaenoic n-3 in total liver. These results demonstrate that in humans or animals that are on the border of EFA deficiency, 1% cholesterol is enough to lead them to a true deficiency due to a further decrease of arachidonic and docosahexaenoic n-3 acids.
EFECTO DEL COLESTEROL DE LA DIETA SOBRE LA BIOSÍNTESIS DE LOS ACIDOS GRASOS NO SATURADOS
(especial para SIIC © Derechos reservados)
Artículo completo
Introducción
Es científicamente conocido, aunque no siempre entendido por el público, que el colesterol es un componente indispensable para el hombre y los mamíferos, con una función primordial en la determinación de las características biofísicas de las membranas plasmáticas celulares. Se lo encuentra en las caveole y rafts y es el sustrato para la biosíntesis de las hormonas esteroides sexuales y corticoides, entre otras. La mayor parte de ese colesterol es biosintetizado en el hígado y otra parte proviene de los alimentos de orígen animal, especialmente huevos. El colesterol es transportado en la sangre por varias lipoproteínas. El hígado lo oxida a ácidos biliares y el exceso es eliminado en esta forma. Los organismos con metabolismo normal no tienen problemas. Pero si se lo altera, por efecto del exceso y principalmente por su oxidación a 7-ß-hidroxicolesterol se producen alteraciones en los macrófagos e íntima de las paredes sanguíneas y problemas cardiovasculares, entre otros. Un exceso de colesterol en la dieta indudablemente produce entonces una acción nociva.Por otro lado, en 19291 se comenzó a conocer que la carencia en la dieta de los ácidos grasos linoleico (n-6) y a-linolénico (n-3) produce trastornos biológicos importantes y progresivos. Luego se demostró que ello se debía fundamentalmente a la disminución de los ácidos poliinsaturados eicosa-8,11,14-trienoico (20:3n-6), araquidónico (20:4n-6) y docosa-4,7,10,13,16-pentenoico (22:5n-6) en el primer caso y eicosa-5,8,11,14,17-pentenoico (20:5n-3) y docosa-4,7,10,13,16,19-hexenoico (22:6n-3) en el segundo. Estos ácidos son sintetizados a partir de aquellos por reacciones de desaturación y elongación sucesivas. De ellas, las reacciones de desaturación catalizadas por la D6 desaturasa,2 y en segundo lugar por la D5 desaturasa, controlan su formación.En líneas generales, algunos de los efectos biológicos que producen los ácidos poliinsaturados 20:3n-6, 20:4n-6 y 22:5n-3 tienen lugar a partir de su transformación a eicosanoides (prostaglandinas, prostaciclina, tromboxanos y leucotrienos), además de la conversión del ácido araquidónico en hepoxilinas y lipoxinas y formación de anandamida.Los ácidos 22:6n-3 y 20:4n-6 tienen una acción importante en el cerebro. La disminución del aporte de 22:6n-3 produce marcada disminución en el desarrollo de la capacidad mental de los niños y una alteración de los fotorreceptores de la retina. Además son componentes de los fosfolípidos de las membranas y pueden regular ciertos factores de transcripción.Ya en 19603,4 se había observado la existencia de algunas relaciones metabólicas entre el colesterol y los ácidos grasos esenciales. Con los medios que se poseía en aquel tiempo, se pudo reconocer que la ingestión de colesterol incrementaba en los animales los efectos biológicos de la deficiencia de ácidos grasos esenciales. Mucho después, desde 1986,5-8 hemos demostrado que una adición de 1% de colesterol a la dieta de las ratas era suficiente para disminuir la actividad de las D6 y D5 desaturasas y aumentar la correspondiente de la D9, disminuyendo la proporción de los ácidos 20:4n-6 y 22:6n-3 en los lípidos. El efecto era reversible. Esos animales, sin embargo, habían ingerido dietas ricas en ácido linoleico y ha sido demostrado que la adición de ese ácido a la dieta disminuye la actividad y el ARNm de las D6 y D5 desaturasas.9,10 Pero pese a ello, el efecto era específico del colesterol.De cualquier manera, desde el punto de vista nutricional resultó importante determinar si el colesterol dietario podía empujar a un organismo alimentado con una dieta pobre en AGE a una real carencia y producir así efectos negativos adicionales.Para resolver ese problema se llevó a cabo una experiencia consistente en alimentar ratas recién destetadas con una dieta pobre en AGE. Luego se le adicionó 1% de colesterol y se estudió el comportamiento de las D9, D6 y D5 desaturasas y los cambios de composición acídica de los lípidos hepáticos. Los resultados demostraron que el efecto era netamente estimulador de una carencia de AGE.Parte experimental
• Productos químicos
Los ácidos esteárico 1-14C (56 mCi/mmol), linoleico 1-14C (55 mCi/mmol) y eicosa-8,11,14-trienoico 1-14C (52 mCi/mmol) fueron provistos por Amersham (Inglaterra). El cADN de la D9 desaturasa y el cADN de la D6 desaturasa fueron donaciones del Dr. Juris Ozols de la Universidad de Conneticut, Farmington, Estados Unidos y el Dr. Tsunehiro Aki de la Universidad de Hiroshima, Hiroshima, Japón, respectivamente.• Animales y dietas
Ratas Wistar machos recién destetadas fueron alimentadas durante 10 días con una dieta control baja en AGE, con 62% de almidón, 22.6% de caseína deslipidizada, 9.4% de manteca de cacao, 2% de vitaminas y 4% de minerales. Sólo 4.5% de los ácidos grasos correspondían al ácido linoleico y 34.0% al oleico. Entonces, mientras un grupo de animales siguió siendo alimentado con esa dieta, otro fue alimentado con la misma dieta más 1% de colesterol. Grupos correspondietes de estos animales fueron sacrificados luego de 1, 3, 7 y 21 días. Los hígados fueron extraídos, homogeneizados y los microsomas separados en la forma corriente.11 Las reacciones se realizaron utilizando microsomas hepáticos.• Técnicas analíticas
Las técnicas analíticas para determinar colesterol, ésteres de colesterol, fósforo, composición de los ácidos grasos, y actividades enzimáticas de las desaturasas microsomales fueron detalladas anteriormente11 En la medición de las desaturasas se usaron los medios radiocromatográficos corrientes de nuestro Instituto.11La determinación de la abundancia de los ARNm de las D9 (SCD-1) y D6 desaturasas por Northern blots fueron también detallados anteriormente.11Resultados
• Efecto del colesterol dietario entre el contenido de colesterol libre y esterificado de hígado
En primer lugar se observó que el colesterol agregado a los alimentos era absorbido e incorporado en el hígado aumentando el contenido total de la víscera. Sin embargo, al estudiar la distribución entre el colesterol libre y el esterificado, se observó que sólo se incrementaba realmente la proporción del esterificado. En los microsomas, los ésteres de colesterol, pero no el colesterol libre, aumentaron ya en la primera hora y luego se mantuvieron en ese tenor. En hígado total el aumento de los ésteres de colesterol fue también inmediato, pero siguió creciendo con el tiempo durante los 21 días (cuadro 1).

• Efecto del colesterol sobre la 9 desaturasa
La actividad enzimática de la desaturación hepática del ácido esteárico a oleico aumentó significativamente por efecto del colesterol, pero sólo se observó a los 7 y 21 días, no antes (figura 1).
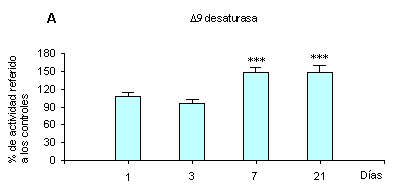

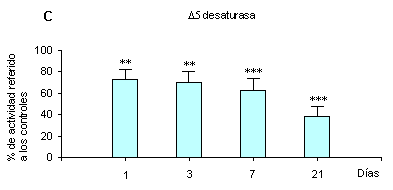
Figura 1. Efecto del colesterol dietario sobre las actividades desaturantes (D9, D6 y D5) de microsomas hepáticos durante un período de 21 días. Los datos representan la media ± DE (n = 4 animales por lote). Significancia estadística respecto a los controles. *** p < 0.001. ** p < 0.01 utilizando test de Student.
En cambio, el incremento relativo del ARNm de la D9 desaturasa hepática (SCD-1) de los animales que recibieron colesterol en relación con los controles se produjo ya a las 24 horas, antes que se manifestara un incremento de la actividad enzimática (figura 2).


Figura 2. Efecto de una dieta rica en colesterol sobre los niveles de ARNm de la estearoil-CoA desaturasa-1 (SCD1) y de la D6 desaturasa hepáticas. A, autorradiografías representativas. B, niveles de ARNm normalizados respecto a los lotes controles.
• Efecto del colesterol sobre las D6 y D5 desaturasas
El colesterol agregado a la dieta provocó, en comparación con los controles, un descenso de las actividades de ambas D6 y D5 desaturasas hepáticas, menos marcado para la D6 desaturasa que para la D5 (figura 1). Estos resultados obtenidos con animales alimentados con una dieta pobre en AGE son similares a los que observamos en las ratas alimentadas con dietas ricas en ácido linoleico (13% de aceite de maíz).5,6Sin embargo, al medir la abundancia del ARNm de la D6 desaturasa hepática por el método del Northern blot, quedó demostrado que la dieta con 1% de colesterol no lo modificó en todos los períodos del experimento hasta los 21 días (figura 2). En consecuencia el efecto depresor del colesterol sobre la actividad de la D6 desaturasa no se produciría a ese nivel.• Efecto del colesterol sobre la composición acídica de los lípidos hepáticos
El efecto del colesterol dietario sobre la composición de los ácidos grasos de los lípidos hepáticos totales y los lípidos microsomales correspondientes se ven claramente en los cuadros 2 y 3. En ellos sólo hemos incluido los valores hallados luego de 1 día y 21 días de dieta. Los correspondientes a 7 días pueden consultarse en la publicación referida anteriormente,11 y sólo señalan la progresión de los valores de 1 día a 21 días.

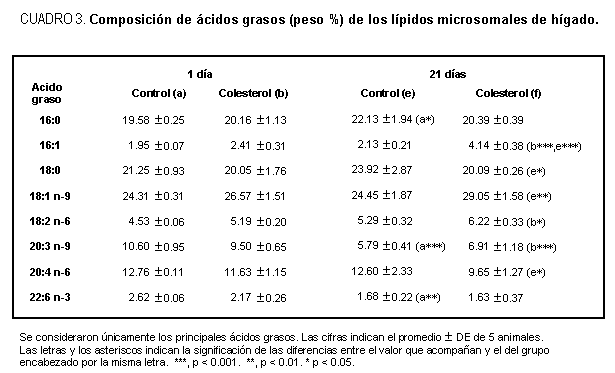
Tanto en los lípidos totales hepáticos como microsomales se observa que la dieta pobre en ácido linoleico provoca en los animales una cierta carencia de AGE debido a los bajos contenidos de ácido linoleico hallados que, sin embargo, se mantienen constantes y similares tanto en los animales control como en los tratados con colesterol. Ese efecto carencial también se manisfiesta por la aparición del ácido eicosa-5,8,11-trienoico (20:3n-9) derivado del ácido oleico.El colesterol provocó una disminución progresiva del ácido araquidónico desde el primer día hasta los 21, tanto en hígado total como es sus microsomas. El descenso de la proporción de 22:6n-3 sólo fue significativo en los lípidos totales hepáticos; sin embargo, hay que señalar que ese ácido, y debido a la composición acídica de la dieta, es sólo un componente minoritario. Los datos concuerdan con el resultado que puede producir sobre la composición lipídica la disminución de la actividad de las D6 y D5 desaturasas, y que también se ha observado por la acción de otros factores moduladores de esas enzimas.12,13Tanto en los lípidos totales del hígado o sus microsomas (cuadros 2 y 3) y en los ésteres de colesterol respectivos11 observamos claramente que el colesterol provoca un incremento progresivo de la proporción de los ácidos palmitoleico y oleico, que sabemos son sintetizados por la D9 desaturasa a partir de los ácidos palmítico y esteárico, respectivamente. Estos datos, en consecuencia, se correlacionan bastante bien con el aumento del ARNm y actividad de la D9 desaturasa hepática observados en las figuras 1 y 2. Además, están de acuerdo con resultados observados anteriormente5,6,14 en animales en los cuales el colesterol se agregó a una dieta rica en ácido linoleico.Discusión
• Incorporación del colesterol dietario en el hígado
Los resultados señalados muestran claramente (cuadro 1) que el colesterol ingerido llega enseguida al hígado y en el retículo endoplásmico no incrementa realmente la proporción de colesterol libre sino la del esterificado, sólo hasta cierto límite. Esta esterificación la realiza la acil-CoA colesterol aciltransferasa (ACAT), enzima localizada en esa organela, que se presenta en los roedores en dos formas ACAT-1 y ACAT-2.15,16 Sin embargo, la enzima realmente activa en hígado sería la ACAT-2. El exceso de ésteres de colesterol es aparentemente removido del retículo endoplásmico; por un lado, pasaría al citosol formando las partículas citosólicas (cytosolic droplets)17 y tornando al colesterol inofensivo para la célula, y por otro lado, pasándolos al torrente sanguíneo bajo la forma de lipoproteínas junto a la apoproteína B y triacilgliceroles.18El incremento progresivo y continuo de la proporción del colesterol esterificado sobre el libre en el hígado total observado en el cuadro 1 por efecto de la incorporación del colesterol a la dieta está de acuerdo con esa interpretación. Además, esos resultados sugieren que la ACAT cumpliría una función importante en la homeostasis del colesterol libre celular.• Efecto sobre la D9 desaturasa
Los resultados presentados en las figuras 1 y 2 claramente señalan, comparados con los de las publicaciones de Leikin y colaboradores5,6 y Landau y colaboradores,14 que tanto en los animales alimentados con una dieta rica en ácido linoleico, que deprime su actividad,19 como con una dieta pobre, que corresponde a este experimento, el agregado de 1% de colesterol en los alimentos aumenta en ambos casos no sólo el ARNm de la D9 desaturasa hepática (SCD-1) sino también su actividad enzimática. En el experimento presente el incremento del ARNm precede al de la actividad enzimática. Es importante señalar que de las dos formas SCD-1 y SCD-2 de la D9 desaturasa halladas hasta ahora en la rata (en la laucha se encontró también una isoforma SCD-3), la forma SCD-1 es típica del hígado y desatura tanto a la palmitoil-CoA como a la estearoil-CoA. Esa isoforma es la que activa el colesterol, según resulta de los experimentos señalados.Es importante además señalar que el incremento de la actividad de la D9 desaturasa hepática que produce el colesterol se adiciona en el presente experimento al que ocasiona ya la dieta básica pobre en ácido linoleico. Este efecto aditivo se manifiesta también en el aumento de los ya altos contenidos de ácido oleico de los lípidos hepáticos totales y microsomales (cuadros 2 y 3) y en los ésteres del colesterol11 al ingerir las ratas el colesterol. Ese aumento de la relación 18:1n-9/18:0 en los fosfolípidos microsomales produce un efecto incrementador de la fluidez de esas membranas que puede acarrear efectos biológicos importantes y compensar, por ejemplo, otros cambios provocados por el descenso en la fosfatidilcolina de las especies moleculares 18:0-20:4 y 16:0-20:4.20Es esperable que el efecto observado del colesterol sobre la 9 desaturasa esté mecanísticamente relacionado con la activación y desactivación de algún sterol regulatory element binding protein (SREBP) que en sus diversas formas 1a, 1c y 2 intervienen una vez truncados como factores de transcripción a nivel nuclear. Sus mecanismos de activación y funciones están en pleno período de investigación, pero se sabe ya que regulan la transcripción de una serie de enzimas sintetizadoras de ácidos grasos, entre ellas las desaturasas.21,22• Efecto sobre las D6 and D5 desaturasas
La depresión que provoca el colesterol de la dieta sobre la actividad de las D6 y D5 desaturasas hepáticas observada en animales alimentados con raciones ricas en ácido linoleico,5,6 se repite en este experimento con raciones pobres en ese ácido esencial (figura 1). Ello demuestra que el efecto depresor del colesterol se observa aún pese al aumento del ARNm de esas enzimas que provoca por sí una dieta pobre en linoleico.9,10El descenso de esas actividades enzimáticas explicarían bien (y consideramos que son la causa fundamental) la disminución de la proporción del ácido araquidónico en los lípidos hepáticos totales y microsomales y del 22:6n-3 en los lípidos hepáticos totales (cuadros 2 y 3) al disminuir su biosíntesis. Ellos son ácidos grasos indispensables y fundamentales. Ello también fue observado5,6 ocurre con las ratas alimentadas con dietas ricas en ácido linoleico. Bernasconi y colaboradores20 además demostraron que el agregado de colesterol a una dieta rica en linoleico deprimía específicamente la proporción de las especies moleculares de fosfatidilcolina microsomal 18:0/20:4n-6 y 16:0/20:4n-6, aumentando las especies que contenían 18:1. Pero ahora adquiere un significado nutricional más importante porque demuestra que animales o seres humanos que están al borde de una carencia de ácidos grasos esenciales pueden caer en ella al ingerir cantidades mayores de colesterol. Este caso puede ser bastante frecuente en pueblos desnutridos, pero menos posible en aquellos que se alimentan con la comúnmente llamada en Estados Unidos western diet, que es muy rica en ácido linoleico de origen vegetal. Por otro lado, la ingestión de alimentos de orígen animal ricos en ácido araquidónico o 22:6n-3, éste último presente en pescados marinos especialmente, compensarían esa carencia a pesar de disminuir aún más la actividad de las D6 y D5 desaturasas.23,24La figura 2 además señala que el efecto depresor del colesterol sobre la D6 desaturasa no se produciría provocando menor abundancia de su ARNm. Ello también aparentemente descartaría una intervención del SREBP en tal efecto porque se ha probado que al menos el SREBP-1c activa a las D6 y D5 desaturasas activando la transcripción de los ARNm de esas desaturasas.22 Hasta ahora, en consecuencia, sólo se deduce que esa modulación la produciría el colesterol aparentemente en un paso posterior al del ARNm. Tampoco hemos podido confirmar la hipótesis vertida por nosotros5,6 anteriormente, que tal regulación se podría producir por alteración de la estructura y propiedades biofísicas de la membrana microsomal.Hasta ahora no hemos determinado cuál es el efecto directo de la dieta con colesterol sobre la abundancia del ARNm de la D5 desaturasa, pero la similitud de la secuencia de los nucleótidos de los genes de ambas D6 y D5 desaturasas,10,25 la existencia de una región común de 11.000 bp para ambos genes10 y la extraordinaria similitud de su regulación en los animales por los diversos factores26 permiten prever que debe ser muy semejante al que ejerce sobre la D6 desaturasa. De cualquier manera los resultados señalan que el mecanismo modulador de la actividad de las D6 y D5 desaturasas por acción del colesterol sería diferente del empleado para la D9 desaturasa.En conclusión, y especialmente debido al efecto depresor del colesterol dietario sobre la actividad de las D6 y D5 desaturasas, aquel disminuye el contenido de los ácidos indispensables araquidónico y docosahexenoico (n-3) en los lípidos. Ello trae con consecuencia, sobre todo en animales y seres humanos que se alimentan con dietas pobres en los ácidos grasos esenciales de origen vegetal linoleico y -linolénico, un estado de verdadera carencia de ácidos grasos esenciales. Este problema puede ser importante, fundamentalmente en los pueblos desnutridos. El efecto se adicionaría así a los problemas que provoca el exceso de colesterol en aquellos pacientes que tienen problemas en su metabolismo.Abreviaturas. AGE, ácidos grasos esenciales. SCD-1, estearoil-CoA desaturasa-1. SREBP, sterol regulatory element binding protein
Bibliografía del artículo
- Burr G.O., and Burr M.M. (1929) A new deficiency disease produced by the rigid exclusion of fat from the diet. J. Biol. Chem. 82:345-367.
- Brenner R.R. (1977) Regulatory function of D6 desaturase key enzyme of polyunsaturated fatty acid synthesis. Adv. Exp. Med. Biol. 83: 85-101.
- Holman R.T., and Peiffer J.J. (1960) Acceleration of essential fatty deficiency by dietary cholesterol. J. Nutr. 70: 411-417.
- Gambal D., and Quackenbush F.W. (1960) Effect of cholesterol and other substances on essential fatty acid deficiencies. J. Nutr. 70: 497-501.
- Leikin A.I., and Brenner R.R. (1987) Cholesterol-induced microsomal changes modulate desaturase activities. Biochim. Biophys. Acta 922: 294-303.
- Leikin A.I., and Brenner R.R. (1988) In vivo cholesterol removal from liver microsomes induces changes in fatty acid desaturase activities. Biochim. Biophys. Acta 963: 311-319.
- Garg M.L., Snowswell A.M., and Sabine J.R. (1986) Influence of dietary cholesterol on desaturase enzymes of rat liver microsomes. Prog. Lipid Res. 25: 639-644.
- Igal R.A., and Gómez Dumm I.N.T. de (1994) Changes in adrenal gland and liver polyunsaturated fatty acids biosynthesis in hypercholesterolemic rats. Nutr. Res. 14: 241-254.
- Nakamura M.T., Cho H.P., and Clarke S.D. (2000) Regulation of hepatic D6 desaturase expression and its role in the polyunsaturated fatty acid inhibition of fatty acid synthase gene expression in mice. J. Nutr. 130: 1561-1565.
- Cho H.P., Nakamura M.T., and Clarke S.D. (1999) Cloning expression and fatty acid regulation of the human D5 desaturase. J. Biol. Chem. 274: 37335-37339.
- Brenner R.R., Bernasconi A.M., González M.S., and Rimoldi O.J. (2002) Dietary cholesterol modulates D6 and D9 desaturase mRNAs and enzymatic activity in rats fed a low-EFA diet. Lipids 37: 375-383.
- Brenner R.R., Bernasconi A.M., and Garda H.A. (2001) Effect of experimental diabetes on the fatty acid composition, molecular species of phosphatidyl-choline and physical properties of hepatic microsomal membranes. Prostaglandins, Leukot. Essent. Fatty Acids 63: 167-176.
- Brenner R.R., Ayala S., and Garda H.A. (2001) Effect of dexamethasone on the fatty acid composition of total liver microsomal lipids and phosphatidyl-choline molecular species. Lipids 36: 1337-1341.
- Landau J.M., Sekowski A., and Hamm M.W. (1997) Dietary cholesterol and the activity of stearoyl-CoA desaturase in rats: Evidence for an indirect effect. Biochim.Biophys. Acta 1345: 349-357.
- Cases S., Novak S., Zheng Y., Myers H.M., Lear S.R., Sandet E., Welch C.B., Luses A.J., Spencer T.A., Krause B.R., Erickson S.K., and Farese R.V. (1998) ACAT-2, Second mammalian acyl-CoA: Cholesterolacyltransferase. J. Biol. Chem. 273: 26755-26764.
- Buhman K.F., Accad M., and Farese R.V.,Jr. (2000) Mammalian acyl-CoA: Cholesterol acyltransferases. Biochim. Biophys. Acta 1529: 142-154.
- Kellner-Weibel G., McHendry-Rinde B., Haynes M.P., and Adelman S. (2001) Evidence that newly synthesized esterified cholesterol is deposited in existing cytoplasmic lipid inclusions. J. Lipid Res. 42: 768-777.
- Davis R. A. (1999) Cell and molecular biology of the assembly and secretion of apolipoprotein B-containing lipoproteins by the liver. Biochim. Biophys.Acta 1440: 1-31.
- Ntambi J.M. (1992) Dietary regulation of stearoyl-CoA desaturase 1 gene expression in mouse liver. J. Biol. Chem. 267: 10925-10930.
- Bernasconi A.M., Garda H.A., and Brenner R.R. (2000) Dietary cholesterol induces changes in molecular species of hepatic microsomal phosphatidyl choline. Lipids 35: 1335-1344.
- Shimomura I., Shimano H., Korn B.S., Bashmakov Y., and Horton J.D. (1998) Nuclear sterol regulatory element-binding proteins activate genes responsible for entire program of unsaturated fatty acid biosynthesis in transgenic mouse liver. J. Biol. Chem. 273:35299-35306.
- Matsuzaka T., Shimano H., Yahagi N., Amemiya-Kudo, M., Yoshikawa T., Hasty A.H., Tamura Y, Osuga J., Okazaki H., Iizuka Y., Takahashi A., Sone H., Gotoda T., Ishibashi S., and Yamada N. (2002) Dual regulation of mouse 5 and 6 desaturase gene expression by SREBP-1 and PPAR-. J. Lipid Res. 43: 107-111.
- Brenner R.R., Peluffo R.O., Nervi A.M., and De Tomás M.E. (1969) Competitive effect of a and g-linolenyl-CoA and arachidonyl-CoA in linoleyl-CoA desaturation to g-linolenyl-CoA. Biochim. Biophys. Acta 176: 420-422.
- Brenner R.R., and Peluffo R.O. (1967) Inhibitory effect of docosa-4,7,10,13,16,19-hexaenoic acid upon the oxidative desaturation of linoleic into g-linolenic acid and the a-linolenic into octadeca-6,9,12,15-tetraenoic acid. Biochim.Biophys. Acta 137: 184-186.
- Zolfaghari R., Cifelli C.J., Banta M.D., and Ross A.C. (2001) Fatty acid D5 desaturase mRNA is regulated by dietary vitamin A and exogenous retinoic acid in liver of adult rats. Arch. Biochem. Biophys. 391: 8-15.
- Brenner R.R. (2002) Hormonal modulation of D6 and D5 desaturases: Case of diabetes. Prostaglandins, Leukot. & Essent. Fatty Acids (En prensa).
©
Está
expresamente prohibida la redistribución y la redifusión de todo o parte de los
contenidos de la Sociedad Iberoamericana de Información Científica (SIIC) S.A. sin
previo y expreso consentimiento de SIIC

