A DESCOBERTA DE VIAS METABOLICAS CELULARES PARTICIPANTES DE PROCESSOS ONCOGENICOS E O DESENVOLVIMENTO DE FARMACOTERAPIAS ESPECIFICAS
(especial para SIIC © Derechos reservados)
Coautores
Greiciane Gaburro Paneto* Francisco De Paula Careta** Beatriz Alves Vianna*** Bruno Marcos Verbeno**** Marcos do Carmo Oyama*****
Farmacêutica, cursando habilitacao em Análises Clínicas. Universidade Federal do Espíritu Santo, Brasil.*
Graduando em Ciências Biológicas. Universidade Fedeal do Espírito Santo, Brasil.**
Graduanda em Ciências Biológicas***
Graduando em Ciências Biológicas.****
Biólogo. Universidade Federal do Espírito Santo, Brasil.*****
Recepción del artículo: 20 de abril, 2004
Aprobación: 0 de , 0000
 Conclusión breve
A investigação de vias metabólicas intracelulares participantes de processos oncogênicos e sua compreensão detalhada podem permitir o desenvolvimento de terapias gênicas ou farmacológicas mais específicas e eficientes no combate ao câncer
Resumen
A investigação de vias metabólicas intracelulares participantes de processos oncogênicos e sua compreensão detalhada podem permitir o desenvolvimento de terapias gênicas ou farmacológicas mais específicas e eficientes no combate ao câncer. A partir de estudos anteriores, utilizando o oncogene GLI, foi possível estabelecer que, ao menos in vitro e antes das células atingirem a confluência de 100%, as células que expressam esse oncogene são sensíveis ao tratamento com o macrolídeo rapamicina. Outros oncogenes, ao contrário, demonstraram induzir focos resistentes a essa droga. Através da utilização de chips de DNA foram identificados vários alvos transcripcionais do oncogene GLI e um deles, o WNT2B, demonstrou ser necessário à transformação tumoral, mas não sendo capaz de causar tumorigênese isoladamente. Os resultados apontam para a utilização de terapias farmacológicas de concepção moderna na cura do câncer visando a potencialização da ação terapêutica e a diminuição dos efeitos colaterais. Avanços tecnológicos, como os chips de DNA e o sequenciamento de SNPs (Single Nucleotide Polymorphism), permitirão a caracterização polimórfica de cada indivíduo e a escolha da melhor estratégia terapêutica individualizada ao paciente.
Palabras clave
Oncogenes, GLI, WNT2B, chips de DNA
Clasificación en siicsalud
Conclusión breve
A investigação de vias metabólicas intracelulares participantes de processos oncogênicos e sua compreensão detalhada podem permitir o desenvolvimento de terapias gênicas ou farmacológicas mais específicas e eficientes no combate ao câncer
Resumen
A investigação de vias metabólicas intracelulares participantes de processos oncogênicos e sua compreensão detalhada podem permitir o desenvolvimento de terapias gênicas ou farmacológicas mais específicas e eficientes no combate ao câncer. A partir de estudos anteriores, utilizando o oncogene GLI, foi possível estabelecer que, ao menos in vitro e antes das células atingirem a confluência de 100%, as células que expressam esse oncogene são sensíveis ao tratamento com o macrolídeo rapamicina. Outros oncogenes, ao contrário, demonstraram induzir focos resistentes a essa droga. Através da utilização de chips de DNA foram identificados vários alvos transcripcionais do oncogene GLI e um deles, o WNT2B, demonstrou ser necessário à transformação tumoral, mas não sendo capaz de causar tumorigênese isoladamente. Os resultados apontam para a utilização de terapias farmacológicas de concepção moderna na cura do câncer visando a potencialização da ação terapêutica e a diminuição dos efeitos colaterais. Avanços tecnológicos, como os chips de DNA e o sequenciamento de SNPs (Single Nucleotide Polymorphism), permitirão a caracterização polimórfica de cada indivíduo e a escolha da melhor estratégia terapêutica individualizada ao paciente.
Palabras clave
Oncogenes, GLI, WNT2B, chips de DNA
Clasificación en siicsalud
Artículos originales> Expertos del Mundo>
página www.siicsalud.com/des/expertos.php/68047
Especialidades
 Principal: Genética Humana, Oncología,
Principal: Genética Humana, Oncología,
Relacionadas: Farmacología, Medicina Interna,
Enviar correspondencia a:
Iuri Drumond Louro. Rua Horacio Andrade de Carvalho, 210, Vitoria, ES, 29052-620, Brasil Drumond Louro, Iúri
THE DISCOVERY OF ONCOGENIC INTRACELLULAR PATHWAYS AND THE DEVELOPMENT OF NOVEL CANCER THERAPIES
Abstract
The investigation of intracellular oncogenic pathways and their further understanding may enable the development of novel pharmacological therapies or gene therapies, specific and effective for cancer treatment. Following earlier studies utilizing the GLI oncogene, it was possible to establish, at least in vitro and before cells reached 100% confluence, that GLI transformed cells were sensitive to macrolide rapamycin. In contrast, other oncogenes showed resistance to this drug. DNA microarrays were used to identify several transcriptional targets of GLI and one of them, WNT2B, was necessary for oncogenic transformation, but was not capable of inducing malignancy on its own. These results suggest the possibility of utilizing new drug therapies in cancer treatment, attempting to maximize therapeutic effectiveness and minimize side effects. Modern techniques, such as DNA Chips and SNP (Single Nucleotide Polymorphism), will probably allow for the polymorphic characterization of every individual and will enable physicians to choose a better therapeutic strategy for each patient.
A DESCOBERTA DE VIAS METABOLICAS CELULARES PARTICIPANTES DE PROCESSOS ONCOGENICOS E O DESENVOLVIMENTO DE FARMACOTERAPIAS ESPECIFICAS
(especial para SIIC © Derechos reservados)
Artículo completo
Introdução
A investigação de vias bioquímicas intracelulares causadoras ou participantes dos processos oncogênicos é de grande valia para a elaboração de estratégias terapêuticas eficientes e específicas no combate ao câncer.
A partir de estudos anteriores,1 estabelecemos que células que expressam o oncogene GLI são sensíveis ao tratamento com o macrolídeo imunossupressor rapamicina, pelo menos in vitro e antes das células atingirem a confluência de 100%. Em experimentos semelhantes, a rapamicina também mostrou-se capaz de sensibilizar focos de células que expressam GLI, enquanto outros oncogenes, como o RAS, induziram focos resistentes à droga (figura 1). Isso sugere que terapias mais específicas podem ser desenvolvidas para cada mecanismo de indução tumoral, tornando o tratamento mais eficaz.
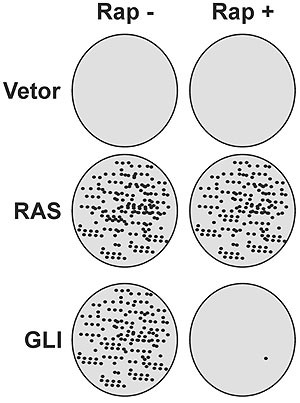
Figura 1. Representação gráfica da inibição do oncogene GLI pelo macrolídeo rapamicina (10 ng/ml). A transformação in vitro só ocorre quando os oncogenes RAS ou GLI são transfectados nas células em cultura. O número de focos neoplásicos obtidos com o oncogene RAS não é afetado pelo tratamento com rapamicina. Quando o mesmo tratamento é aplicado ao oncogene GLI ocorre uma drástica inibição do número de focos tumorais.
A análise da expressão gênica pode ser realizada através de métodos específicos como os chips de DNA, que são métodos altamente computadorizados, permitindo a análise de expressão de milhares de genes simultaneamente e possibilitando a caracterização do padrão de expressão gênica de cada célula.
A sensibilidade provocada pelo GLI à rapamicina parece ser devida a uma incapacidade destas células de induzirem a tradução protéica na presença dessa droga (figura 2).
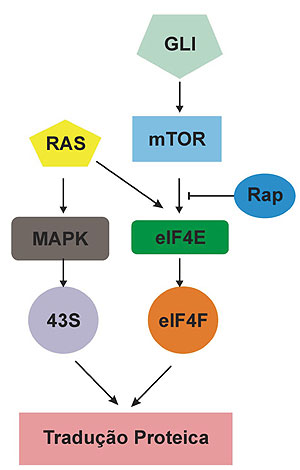
Figura 2. Utilização de vias metabólicas distintas por oncogenes diferentes. A regulação do início da tradução protéica pode ser feita de mais de uma maneira. O oncogene RAS é capaz de promover a tradução protéica mesmo na presença da rapamicina, indicando a utilização de uma via alternativa. Enquanto o oncogene GLI depende exclusivamente da via do eIF4F para induzir a tradução protéica.
Para identificar mensagens induzidas por este oncogene utilizamos um método baseado na amplificação seletiva por PCR de genes expressos em células que continham apenas o oncogene GLI.1 Contudo, as mensagens inicialmente isoladas se mostraram induzidas somente após muitos dias de transformação oncogênica e foram consideradas de pouca relevância no contexto de eventos iniciais do processo oncogênico. Visando identificar mudanças precoces na expressão gênica após o início da transformação neoplásica, células RK3E foram transformadas com um determinado oncogene e testadas após 2 ou 10 dias, sendo a análise de expressão gênica feita pelo método de PCR seletivo (SSH) ou por chips de DNA. Dessa forma, novos genes induzidos pelo GLI foram identificados. Após a análise qualitativa dos resultados obtidos, algumas vias foram consideradas como possíveis mediadoras da tumorigenicidade do GLI e constituem fortes alvos à terapia gênica ou farmacológica, isoladas ou em combinação.

Figura 3. A via metabólica do WNT-APC-β-Catenina. Via oncogênica clássica, que se inicia no meio extracelular com o ligante WNT e se propaga para o citoplasma através dos participantes representados acima. Na ausência da ativação da via, a β-Catenina se encontra associada ao APC, Axin e GSK3 e portanto inativa. Diversos estímulos, incluindo o WNT, são capazes de induzir a dissociação da β-Catenina deste complexo e o transporte da mesma para dentro do núcleo celular onde, associada aos fatores de transcrição TCF4 e ao cofator CBP, é capaz de induzir a transcrição de genes alvo. Na presença de um alelo mutante do TCF4, TCF4-DN, esta associação intranuclear não resulta num complexo capaz de induzir a transcrição gênica.
Decidimos investigar o gene WNT2B devido a sua participação numa via oncogênica clássica em vertebrados2-6 e por ser induzido por um gene GLI-homólogo em invertebrados (Cubitus interruptus).7-9 Para testar a necessidade da via metabólica de WNT2B no potencial oncogênico de GLI, foi necessário a avaliação da via metabólica dos WNTs (figura 3) e a seleção de um alelo dominante negativo TCF4-DN capaz de abolir a mensagem proveniente do ligante extracelular WNT cujo sinal se propaga para o citoplasma e atinge o núcleo estimulando a expressão gênica. O mutante TCF4-DN foi transfectado por retrovírus em células RK3E, e as células recipientes foram selecionadas com G418, estabelecendo assim a uma linhagem celular que expressa constitutivamente a proteína TCF4-DN e que conseqüentemente abole a via de ativação dos WNTs, também conhecida como via do APC. A seguir, clonamos o gene WNT2B num vetor de expressão retroviral que foi usado para transfectar a linhagem RK3E/TFN2-DN. Após vários dias de cultura, os resultados mostrados na figura 4 demonstram claramente que a terapia gênica utilizada in vitro é capaz de inibir significantemente a transformação pelo GLI, estabelecendo que a via dos WNTs é necessária para a ação neoplásica de GLI.
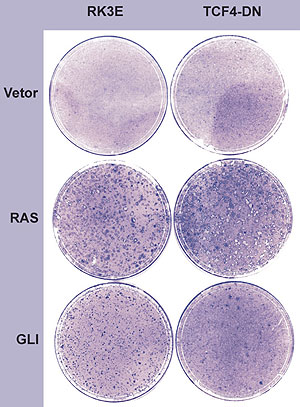
Figura 4. Terapia Gênica in vitro. O bloqueio da transformação maligna induzida pelo GLI é mediado pela expressão do alelo dominante negativo TCF4-DN. O oncogene RAS se mostrou resistente à expressão deste alelo, mas o oncogene GLI demonstrou uma drástica diminuição no número de focos neoplásicos, quando a linhagem receptora expressava constitutivamente este produto gênico inativo e dominante. Isso demonstra a necessidade da atividade da via do WNT-APC-β-Catenina para a transformação maligna pelo oncogene GLI.
A seguir, questionamos se WNT2B seria, por si só, suficiente para induzir a formação de focos tumorais em cultura. Para testarmos essa possibilidade, utilizamos o mesmo vetor de expressão retroviral introduzido em células RK3E. Nesse procedimento, não houve a formação de focos tumorais, determinado assim que essa via é necessária para a ação de GLI, mas não é suficiente para que sozinho induza um processo neoplásico em células RK3E.
Para caracterizar ainda em mais detalhes os genes que são induzidos pelo GLI, uma versão desse gene foi clonada contendo uma porção C-terminal responsiva ao farmacoterápico tamoxifeno. Essa é uma porção modificada do receptor de estrogênio que contém o sítio de ligação com proteínas citoplasmáticas, mas que não possuem qualquer sítio de ligação com o DNA ou de transativação de expressão gênica. A utilização dessa proteína clonada permite que o produto do gene GLI fusionado permaneça inativo no citoplasma, até que se adicione o agente tamoxifeno, quando a proteína é liberada para atingir o núcleo e exercer suas funções de fator de transcrição.10-11 Com esse controle preciso da ativação de GLI fornecido pelo inibidor de síntese proteica ciclohexamida, é possível estudar o mecanismo da expressão gênica passado apenas minutos ou horas, assim como saber se determinado gene é induzido diretamente pelo GLI. Os resultados deste experimento demonstram que GLI não induz diretamente WNT2B, o que seria realizado provavelmente por um de seus genes localizados abaixo de sua cadeia de expressão gênica (figura 5).
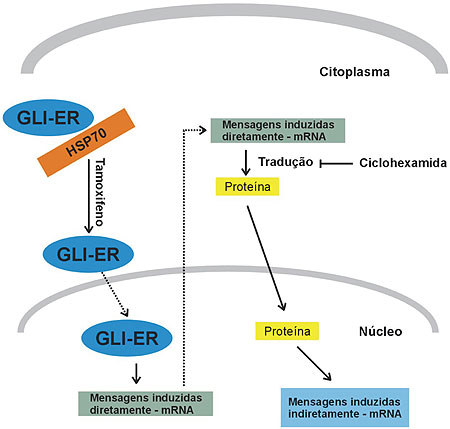
Figura 5. Identificação de mensagens diretamente induzidas pelo GLI (GLI-ER). A porção ER mantém o GLI associado a proteínas citoplasmáticas. Quando o meio de cultura é tratado com tamoxifeno, o GLI-ER se dissocia deste complexo e se localiza no núcleo, onde promove a expressão dos genes diretamente por ele induzidos. Estes, no citoplasma são traduzidos em proteínas e, no caso de serem fatores de transcrição, podem voltar ao núcleo e promover nova transcrição gênica. Esta segunda etapa de transcrição não ocorre na presença do inibidor de tradução protéica ciclohexamida.
Materiais e métodos
Contrução de clones e transdução retroviral: O oncogene GLI, contendo um epítopo de hemaglutinina (HA) na sua porção N-terminal foi clonado no vetor pLJD (MMLV). A transfecção de pLJD-GLI em células RK3E foi previalmente descrita,1 e produziu focos de células malignas que expressavam GLI, de acordo com experimentos de imunoprecipitação com anticorpos 12CA5 (anti-HA). As células transduzidas com LJD ou LJD-GLI foram selecionadas em 400 mg/ml G418 por 2 ou 10 dias. Pelo menos 1 mg de RNA total foi isolado de culturas celulares a uma confluência de aproximadamente 80%, e RNA poliA+ foi selecionado utilizando colunas de oligo-dT celulose.
Chip de DNA: A hibridização, escaneamento e análise dos dados foram realizados utilizando chips de alta densidade (U34A e U34B) e o software GeneChip versão 3.3 (Affymetrix, Santa Clara, California). Os experimentos foram executados em duplicata, em meses diferentes e após transduções retrovirais distintas. 30 mg de cRNA biotinilado e fragmentado foram adicionadas a 0.2 ml de solução de hibridização (0.1M 2-[N-Morpholino] ethanesulfonic acid [MES] pH 6.5-6.7, 1M NaCl, 20 mM EDTA, and 0.01% tween 20). Para controle de qualidade, cada amostra foi previamente hibridizada e analizada em chips TEST ou TEST2 e somente quando todos os controles de qualidade recomendados pelo fabricante se mostraram satisfatórios, a amostra foi hibridizada aos chips U34A e U34B, consecutivamente. Cerca de 40-50% de todos os genes representados dos chips foram considerados presentes e somente os genes alterados em mais de 3X foram utilizados em nossa análise.
Hibridização Subtrativa Supressiva: (Suppression Subtractive Hybridization ou SSH): SSH foi realizada utilizando o kit PCR-SelectTM (Clonetec, Palo Alto, California), contendo modificações determinadas pelo autor e disponíveis ao leitor quando requisitadas. 120 clones foram analizados por Northern Blot, comparando o RNA de células transformadas pelo GLI ou o vetor controle. A seqüência completa dos clones foi determinada e analizada online no Genebank. As seqüências novas foram submetidas ao Genebank para inclusão no banco de dados.
Northern Blots: Os genes induzidos pelo GLI identificados foram isolados através de RT-PCR a partir do RNA de células RK3E-GLI e utilizados como sondas para a hibridização em Northern Blots, seguindo a técnica descrita anteriormente.1Discussão e Conclusão
A descoberta que um membro da via dos WNTs, o WNT2B, é ao mesmo tempo induzido pelo oncogene GLI é um resultado intrigante. A via dos WNTs, também chamada via do APC (Adenomatous Poliposis Coli), constitui uma via oncogênica clássica que participa da formação de pólipos intestinais e tumores deles provenientes.12-16 O potencial neoplásico do GLI foi significativamente reduzido quando a via dos APC foi abolida, ficando demostrado que sua transformação tumoral é dependente desta via. De forma isolada, entretanto, a via do APC não foi capaz de causar transformação maligna nas condições do experimento.
Foi possível demonstrar também, a importância de tecnologias mais modernas como a dos Chips de DNA na detecção da expressão gênica diferencial. Além do GLI, outros oncogenes foram testados em nosso laboratório e os resultados foram animadores observando a especificidade encontrada para cada via metabólica estudada.
É grande a expectativa que os avanços tecnológicos possam facilitar a cura de diversas doenças, e dentre elas o câncer, entretanto, o desenvolvimento científico é paulatino. A terapia gênica, esperança atual da terapia moderna, se defronta com múltiplos obstáculos, como por exemplo a especificidade dos vetores e a duração da expressão gênica, como também a resposta imunológica contra antígenos dos vetores ou proteínas por eles expressas. Nesse contexto, a terapia farmacológica nos parece mais próxima para a obtenção de resultados contra o câncer, devendo, entretanto, ser aplicada de forma modernizada, levando-se em consideração a minimização dos efeitos colaterais e a potencialização do efeito terapêutico. Os mesmos avanços tecnológicos permitirão a caracterização polimórfica de cada indivíduo (sequenciamento de SNPs) e a sua susceptibilidade ao fármaco específico, permitindo a escolha da terapêutica ideal evitando a exposição indevida do paciente ao risco de reações adversas e efeitos colaterais.
Los autores no manifiestan conflictos.
Bibliografía del artículo
- Louro ID, Louro ASP, Lima JLO. Identificação de Mensagens Induzidas por Oncogenes. Rev Soc Bras de Cancerol 1999; 7: 30-39.
- Sunaga N, Kohno T, Kollings FT, et al. Constitutive activation of the Wnt signaling pathway by CTNNB1 (beta-catenin) mutations in a subset of lung adenocarcinoma. Genes Chromosomes Cancer 2001; 30: 316-321.
- Barker N, Clevers H. Catenins, Wnt signaling and cancer.Bioessays 2000; 22: 961-965.
- Sakanaka C, Sun TQ, Williams LT. New steps in the Wnt/beta-catenin signal transduction pathway. Recent Prog Horm Res 2000; 55: 225-236.
- Smalley MJ, Dale TC. Wnt signalling in mammalian development and cancer. Cancer Metastasis Rev 1999; 18: 215-230.
- Porfiri E, Rubinfeld B, Albert I, et al. Induction of a-catenin-LEF-1 complex by wnt-1 and transforming mutants of -catenin. Oncogene 1997; 15: 2833-2839.
- Methot N, Basler K. Suppressor of fused opposes hedgehog signal transduction by impeding nuclear accumulation of the activator form of Cubitus interruptus. Development 2001; 128: 733-742.
- Von Ohlen T, Hooper JE. Hedgehog signaling regulates transcription through Gli/Ci binding sites in the wingless enhancer. Mech Dev 1997; 68: 149-156.
- Aza-Blanc P, Kornberg TB. Ci: a complex transducer of the hedgehog signal. Trends Genet 1999; 15: 458-462.
- Danielian PS, Muccino D, Rowitch DH, et al. Modification of gene activity in mouse embryos in utero by a tamoxifen-inducible form of Cre recombinase. Curr Biol 1998; 8: 1323-1326.
- Hirst SK, Grandori C. Differential activity of conditional MYC and its variant MYC-S in human mortal fibroblasts. Oncogene 2000; 19: 5189-5197.
- Sparks B, Morin PJ, Vogelstein B, et al. Mutational analysis of the APC/beta-catenin/Tcf pathway in colorectal cancer. Cancer Res 1998; 58: 1130-1134.
- Morin PJ, Sparks AB, Korinek V, et al. Activation of Beta-Catenin-Tcf Signaling in Colon Cancer by Mutations in Beta-Catenin or APC. Science 1997; 275: 1787-1790.
- Korinek V, Barker N, Morin PJ, et al. Constitutive transcriptional activation by a b-catenin-Tcf complex in APC -/- colon carcinoma. Science 1997; 275: 1784-1787.
- Su LK, Vogelstein B, Kinzler KW. Association of the APC tumor-suppressor protein with catenins. Science 1993; 262: 1734-1737.
- van der Luijt RB, Tops CM, Vasen HF. From gene to disease; the APC gene and familial adenomatous polyposis coli. Ned Tijdschr Genneskd 2000; 144: 2007-2009.
©
Está
expresamente prohibida la redistribución y la redifusión de todo o parte de los
contenidos de la Sociedad Iberoamericana de Información Científica (SIIC) S.A. sin
previo y expreso consentimiento de SIIC


