Bibliografía del artículo
Bibliografía del artículo
1. Abdella N, Arouj M Al, Nakhi A Al, et al. Non-insulin-dependent diabetes in Kuwait:prevalence rates and associated risk factors. Diabetes Research and Clinical Practice 1998; 42:187-196.
2. Sharma JN. The tissue kallikrein-kininogen-kinin pathways:role in cardiovascular system. Arch Med Res 2005; in press.
3. Sharma JN. Does the kinin system mediate in cardiovascular abnormalities? An overview. J Clin Pharmacol 2003; 43:1187-95.
4. Sharma JN, Sharma J. Cardiovascular properties of the kallikrein-kinin system. Curr Med Res Opin 2002; 18:10-17.
5. Sharma JN. Contribution of kinin system to the antihypertensive action of angiotensin converting enzyme inhibitors. Adv Exp Med Biol 1989; 247A:197-205.
6. Sharma JN. Role of tissue kallikrein-kininogen-kinin pathways in the cardiovascular system. Archives of Medical Research 2006; 37(3):299-306.
7. Sharma JN, Uma K. Effect of captopril on urinary kallikrein, blood pressure and myocardial hypertrophy in diabetic spontaneously hypertensive rats. Pharmacology 2002; 64:196-200.
8. Sharma JN. Involvement of the kinin-forming system in physiopathology of rheumatoid inflammation. Agents Actions 1992; 38 (III):343-361.
9. Sharma JN, Buchanan WW. Pathogenic responses of bradykinin system in chronic inflammatory rheumatoid disease. Exp Toxicol Pathol 1994; 46:421-433.
10. Nagayasu T, Nagasawa S. Studies of human kininogen. Isolation, characterization, and cleavage by plasma kallikrein of high molecular weight (HMW) kininogen. J Biochem 1979; 85:249-258.
11. Leeb-Lundberg LM, Marceau F, Muller-Esterl W, et al. Classification of the kinin receptor family:from molecular mechanisms to pathophysiological consequences. Pharmacol Rev 2005; 57(1):27-77.
12. Sharma JN. Therapeutic prospects of bradykinin antagonists. Gen Pharmacol 1993; 24:267-274.
13. Bhoola KD, Figueroa CD, Worthy K. Bioregulation of kinins: kallikreins, kininogens, and kininases. Pharmacol Rev 1992; 44(1):1-80.
14. Moreau ME, Garbacki N, Molinaro G, et al. The kallikrein-kinin system:current and future pharmacological targets. J Pharmacol Sci 2005; 99:6-38.
15. Weiss AS, Gallin JL, Kaplan AP. Flecher factor deficiency:a diminished rate of Hageman factor activation caused by absence of prekallikrein with abnormalities of coagulation, fibrinolysis, chemotactic activity and kinin generation. J Clin Invest 1974; 53:622-633.
16. Cochrane CG, Revak SD, Wuepper KD. Activation of Hageman factor in solid liquid faces. A critical role of kallikrein. J Exp Med 1973; 138:1564-1583.
17. Motta G, Rojkjaer R, Hasan AAK et al. High molecular weight kininogens regulates prekallikrein assembly and activation on endothelial cells:a novel mechanism for contact activation. Blood 1998; 91:516-528.
18. Shariat-Madar Z, Mahdi F, Schmaier AH et al. Identification and characterization of prolylcarboxypeptidase as an endothelial cell prekallikrein activator. J Biol Chem 2002; 277:I7962-I7969.
19. Griffin JH, Cochrane CG. Human factor XII (Hageman factor). Methods Enzymol 1976; 45:56-65.
20. Thompson RE, Mandle R Jr, Kaplan AP. Association of factor XI and high molecular weight kininogen in human plasma. J Clin Invest 1977; 60(6):1376-80.
21. Silverberg M, Diehl SV. The autoactivation of factor XII (Hageman factor) induced by low-Mr heparin and dextran sulphate. The effect of the Mr of the activating polyanion. Biochem J 1987; 248(3):715-20.
22. Woolly-Miller C, Chao J, Chao L. Restriction fragment length polymorphism's mapped in spontaneously hypertensive rats using kallikrein probs. J Hypertension 1989; 7:865-871.
23. Pravence M, Ken V, Kunes J. Cosegregation of blood pressure with kallikrein gene family polymorphism. Hypertension 1991; 17:242-246.
24. Erdos EG. Angiotensin I converting enzyme and the changes in our concepts through the years. Lewis K Dhal memorial lecture. Hypertension; 1990; 16(4):363-370.
25. Farmer SG, Burch RM. Biochemical and molecular pharmacology of kinin receptors. Ann Rev Pharmacol Toxicol 1992; 32:511-536.
26. Burch RM. Kinin signal transduction:role of phosphoinositides and eicosanoids. J Cardiov Pharmacol 1990; 15 (Suppl. 6):S44-S46.
27. Akbar A, Sharma JN, Yusof APM. Potentiation of bradykinin-induced responses in the intact and denuded epithelium of guinea pig tracheal preparations. Tissue Reactions 1998; XX:95-100.
28. Schini VB, Boulanger C, Regol D et al. Bradykinin stimulates the production of cyclicGMP via activation of B2 receptors in cultured porcine aortic endothelial cells. J Pharmacol Expl Ther 1990; 43:1823-1827.
29. Jaffa AA, Miller DH, Bailey GS et al. Abnormal regulation of renal kallikrein in experimental diabetes. J Clin Invest 1987; 80:1651-1659.
30. Jaffa AA, Rust PF, Mayfield RK. Kinin, a mediator of diabetes induced glomerular hyper filtration. Diabetes 1995; 44:156-160.
31. Harvey JN, Jaffa AA, Margolius HS. Renal kallikrein abnormalities of diabetic kidney. Diabetes 1990; 39:299-303.
32. Platts JK, Meadows P, Harvey JN. The relationship between urinary kallikrein and glomerular filtration rate (GFR) in type-1 diabetes: studies with lithium. Immunopharmacol 1996; 33:351-353.
33. Tschope G, Gavriluk V, Reinecke A. Bradykinin excretion is increased in severely hyperglycemic streptozotocin-diabetic rats. Immunopharmacol 1996; 33:344-348.
34. Vieira MAR, Moreira FM, Maack T. Conversion of T-kinin to bradykinin by the rat kidney. Biochem Pharmacol 1994; 47:1693-1699.
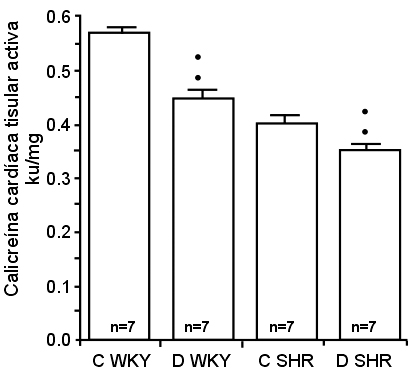
35. Sharma JN, Uma K. Cardiac kallikrein in hypertensive and normotensive rats with and without diabetes. Immunopharmacology 1996; 33:341-343.
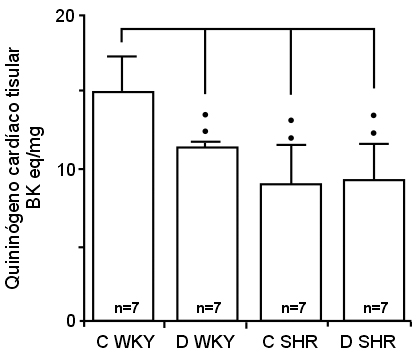
36. Sharma JN, Uma K, Yusof APM. Left ventricular hypertrophy and its relation to cardiac kinin-forming system in hypertensive and diabetic rats. Int J Cardiol 1998; 63:229-235.
37. Sharma JN, Uma K, Yusof APM. Altered cardiac tissue and plasma kininogen levels in hypertensive and diabetic rats. Immunopharmacol 1999; 43:129-132.
38. Tshope C, Reinecke A, Seidl U et al. Functional, biochemical, and molecular investigations of renal kallikrein-kinin system in diabetic rats. Am J Physiol 1999; 277:H2333-2340.
39. Jaffa AA, Durazo-Arvizu R, Zheng D et al. Plasma prekallikrein:a risk marker for hypertension and nephropathy in type 1 diabetes. Diabetes 2003; 52(5):1215-21.
40. Nolly HL, Britos J. Kinin-forming enzyme in rat cardiac tissue. Amer J Physiol 1981; 265:H-1209-H1214
41. Nolly HL, Carretero OA, Sclicli AG. Kallikrein release by vascular tissue. Amer J Physiol 1993; 265:H1209-H1214.
42. Vegh A, Szekeres L, Parratt RJ. Local intracoronary infusions of bradykinin profoundly reduce the severity of ischemia-induced arrhythmia in anaesthetized dogs. Brit J Pharmacol 1991; 104:294-295.
43. Linz W, Wiemer G, Scholkens BA. Bradykinin prevents left ventricular hypertrophy in rats. J Hypertension 1993; 11(Suppl. 5):S96-S97.
44. Vegh A, Rapp JG, Parratt JR. Attenuation of the antiarrhythmic effects of ischemia preconditioning by blocked of bradykinin B2 receptors. Brit J Pharmacol 1994; 107:1167-1172.
45. Walls TM, Sheehy R, Hartman JC. Role of bradykinin in myocardial preconditioning. J Pharmacol Exp Ther 1994; 270:681-689.
46. Sharma JN, Fernandez PG, Laher I. Differential sensitivity of Dahl salt-sensitive and salt-resistant rats to the hypotensive action of acute nifedipine administration. Canad J Physiol Pharmacol 1984a; 62:241-243.
47. Sharma JN. Interrelationship between the kallikrein-kinin system and hypertension: a review. Gen Pharmacol 1988; 19(2):177-87.
48. Sharma JN. The kinin system and prostaglandins in the intestine. Pharmacol Toxicol 1988; 63(5):310-6.
49. Sharma JN. Does kinin mediate the hypotensive action of angiotensin converting enzyme (ACE) inhibitors? Gen Pharmacol 1990; 21(4):451-457.
50. Sharma JN, Uma K, Noor AR. Blood pressure regulation by the kallikrein-kinin system. Gen Pharmacol 1996; 27:55-63.
51. De Freitas FM, Farraco EZ, De Azevedo DF. General circulatory alterations induced by intravenous infusion of synthetic bradykinin in man. Circulation 1964; 29:66-70.
52. Adetuyibi A, Mills IH. Relationship between urinary kallikrein and renal function, hypertension, and excretion of sodium and water in man. Lancet 1972; 2:203-207.
53. Mills IH. The renal kallikrein-kinin system and sodium excretion. Quart J Exp Physiol 1982; 23:175-180.
54. Webster ME, Gilmore JP. Influence of kallidin-10 on renal function. Amer J Physiol 1964; 206:714-718.
55. McGiff JC, Itskovitz HD. Terrango NA. The action of bradykinin and eledoicin in the canine isolated kidney:a relationship to prostaglandins. Clin Sci Molec Med 1975; 49:125-131.
56. Margolius HS, Geller R, Pisano JJ. Altered urinary kallikrein excretion in human hypertension. Lancet 1971; 2:1063-1065.
57. Margolius HS, Geller R, DeJong W. Altered urinary kallikrein excretion in rats. Hypertension Cir Res 1972; 30:358-362.
58. Margolius HS, Horwwitz D, Pisano JJ. Urinary kallikrein excretion in hypertensive man:relationship to sodium intake and sodium-retaining steroids. Circ Res 1974; 35:820-825.
59. Almeida FA, Stella RCR, Voos A. Malignant hypertension:a syndrome associated with low plasma kininogen and kinin potentiating factor. Hypertension 1981; 3:46-50.
60. Sharma JN, Zeitlin IJ. Altered plasma kininogen in clinical hypertension. Lancet 1981; 1:125-126.
61. James FW, Donaldson VH. Decreased exercise tolerance and hypertension in severe hereditary deficiency of plasma kininogens. Lancet 1981; 1:889.
62. Mohsin SSJ, Majima M, Katori M. Important suppressive roles of the kallikrein-kinin system during the developmental stage of hypertension in spontaneously hypertensive rats. Asia Pacific J Pharmacol 1992; 7:73-82.
63. Katori M, Majima M. Role of the renal kallikrein-kinin system in the development of hypertension. Immunopharmacol 1997; 36:237-242.
64. Wang C, Chao L, Chao J. Human tissue kallikrein induces hypotension in transgenic mice. Hypertension 1994; 23:236-243.
65. Sharma JN, Amrah SS, Noor AR. Suppression of hypotensive responses of captopril and enalapril by kallikrein inhibitor aprotinin in spontaneously hypertensive rats. Pharmacology 1995; 50:363-369.
66. Chao J, Chao L. Kallikrein gene therapy in hypertension, cardiovascular and renal diseases. Gen Ther Mol Biol 1998; 1:301-308.
67. Antonacio M. Angiotensin converting enzyme (ACE) inhibitors. Annu Rev Pharmacol Toxicol 1982; 22:57-87.
68. Silberbauer K, Stanek B, Temple H. Acute hypotensive effect of captopril in man modified by prostaglandin synthesis inhibition. Brit J Clin Pharmacol 1982; 14:87S-93S.
69. Sharma JN, Fernandez PG, Kim BK. Cardiac regression and blood pressure control in Dahl rats treated with enalapril maleate (MK 421), an angiotensin converting enzyme inhibitor. J Hypertension 1983; 1:251-256.
70. Edery H, Rosenthal T, Amitzur G. The influence of SQ 20881 on the blood kinin system of renal hypertensive patients. Drug Exp Clin Res 1981; VII:749-756.
71. Sharma JN, Fernandez PG, Kim BK. Systolic blood pressure responses to enalapril maleate (MK 421), an angiotensin converting enzyme inhibitor, and hydrochlorothiazide in conscious Dahl salt-sensitive (S) and salt-resistant (R) rats. Canad J Physiol Pharmacol 1984b; 62:846-849.
72. Smith C, Campbell S, Albano J. Urinary kallikrein excretion in normotensive and hypertensive pregnancies: 8 years later. Immunopharmacol 1999; 44:177-182.
73. Regoli D. Neurohumoral regulation of precapillary vessels: the kallikrein-kinin system. J Cardiovasc Pharmacol 1984; 6 (suppl. 3):S401-S412.
74. Sharma JN, Stewart JM, Mohsin SS et al. Influence of a kinin antagonist on acute hypotensive responses induced by bradykinin and captopril in spontaneously hypertensive rats. Agents Actions Suppl 1992; 38:258-69.
75. Braunwald E. Cardiovascular medicine a turn of the millennium:triumphs, concern and opportunities. New Eng J Med 1997; 337:1360-1369.
76. Locherner W, Parratt JR. A comparison of the effects of locally and systemically administration of kinin on coronary blood flow and myocardial metabolism. Br J Pharmacol Chemother 1966; 26:17-26.
77. Linz W, Wiemer G, Scholkens BA. Contribution of kinins to the cardiovascular action of converting-enzyme inhibitors. Pharmacol Rev 1995; 47:25-50.
78. Zhu P, Zugga CE, Simper D. Bradykinin improves post-ischemic recovery in that rat heart:role of high energy phosphate, nitric oxide and prostacyclin. Carnivals Res 1995; 29:658-663.
79. Madeddu P, Milia AF, Salis MB. Renovascular hypertension in bradykinin B2 receptor knockout mice. Hypertension 1998; 23:503-509.
80. Kichuck MR, Seyedi N, Zhang X. Regulation of nitric oxide production in human coronary micro vessels and the contribution of local kinin formation. Circulation 1996; 94:44-51.
81. Cheng CP, Onishi K, Ohte N. Functional effects of endogenous bradykinin in congestive heart failure. Amer J Coll Cardiol 1998; 31:1679-1686.
82. Koide A, Zeitlin IJ, Parratt JR. Kinin formation in ischemic heart and aorta of anaesthetized rats. J Physiol 1993; 467:125P.
83. Linz W, Wiemer G, Scholkens BA. Bradykinin prevents left ventricular hypertrophy in rats. J Hypertension 1993; 11(Suppl. 5):S96-S97.
84. Hashimto K, Hamamoto H, Honda Y. Changes in components of the kinin system and hemodynamics in acute myocardial infarction. Amer Heart J 1978; 95:619-626.
85. Scholkens BA. Kinins in the cardiovascular system. Immunopharmacol 1996; 33:209-217.
86. Rubin LE, Levi R. Protective role of bradykinin in cardiac anaphylaxis. Circ Res 1995; 76:434-440.
87. Abbas SA, Sharma JN, Yusof APM. Effect of bradykinin and its antagonist on survival time after coronary artery occlusion in rats. General Pharmacol 1999a; 33:243-247.
88. Abbas SA, Sharma JN, Yusof APM. The effect of bradykinin and its antagonist on survival time after coronary artery occlusion in hypertensive rats. Immunopharmacol 1999b; 44:93-98.