Volumen 9, Número 3, Agosto 2003
|
|
Volumen 9, Número 3, Agosto 2003 |
|
Expertos Invitados |
| |
|
|
|
Administración de insulina A menudo los médicos no prestan la suficiente atención a la utilización de una técnica adecuada para la inyección de insulina. Se sabe que el sitio de inyección es una variable importante. La insulina regular aplicada en el abdomen produce disminución en los niveles de glucosa en forma más rápida y de mayor magnitud que la administrada en el muslo. 2 Los dispositivos prellenados de insulina (lapiceras) permiten la medición apropiada de incrementos de una unidad y son particularmente útiles para la administración de pequeńas cantidades de insulina; 3 sin embargo, deben ser usados correctamente. La variabilidad intrasujeto de la insulina NPH es alta cuando se aplica sin la resuspensión que se logra al inclinar de arriba hacia abajo la jeringa o la lapicera unas 10 a 20 veces. Esto es particularmente cierto con el uso de lapiceras, porque la resuspención de la insulina NPH con estos dispositivos toma mayor tiempo. La NPH inyectada sin resuspensión disminuye con la aguja en posición hacia arriba, mientras que la inyección con la aguja hacia abajo, reduce la cantidad de glucosa requerida para mantener la euglucemia para una dosis dada. Además, la inyección con la aguja hacia arriba disminuye en 37% y la aguja hacia abajo aumenta en 59% los efectos biológicos de la NPH observados cuando es administrada completamente mezclada. 4 La técnica de inyección de insulina es un problema entre los usuarios de lapiceras, con 38% de los pacientes que resuspenden de modo inadecuado la insulina o remueven la aguja demasiado pronto, aunque esto no se asoció con una diferencia significativa en la HbA1c (hemoglobina glucosilada) o en los requerimientos de dosis de insulina. 5 Cuando se compararon las técnicas de inyección a 90ş y 40-60ş dentro un pliegue sobreelevado de piel y a 90ş y 40-60ş en forma directa, por medio de la ecografía para rastrear las inyecciones de aire ambiental con agujas de 12 mm de longitud, se observó que 16% fueron intramusculares. La técnica óptima con una aguja de 6 mm es a 90ş dentro de un pliegue elevado de piel ya que logra 98% de inyecciones subcutáneas, mientras que con una aguja de 12 mm esto se consigue con el 94% de las dosis administradas en un ángulo de 40-60ş dentro de un pliegue elevado de piel. 6 Terapia con bomba de insulina Terapia con bombas implantables Hipoglucemia Los rińones tendrían un papel importante ya que los diabéticos tipo 1 muestran menores respuestas a la hipoglucemia del glucagon, norepinefrina y epinefrina y fallas en aumentar la producción renal de glucosa, contrario a lo que ocurre en los controles, quienes evidencian incrementos compensatorios en la producción renal y total de glucosa de mayor magnitud. 15 El uso de un bocadillo nocturno, un enfoque que respeta los horarios para evitar la hipoglucemia nocturna, es efectivo en las personas con diabetes tipo 1. Los niveles de glucemia al momento de acostarse que exceden los 180 mg/dl se asocian en forma infrecuente con hipoglucemia, pero con valores más bajos, la utilización de un aperitivo que contenga carbohidratos o proteínas disminuye en forma efectiva la probabilidad de tales episodios. 16 Si se comparan los efectos de la intoxicación alcohólica leve, la hipoglucemia leve o la combinación de ambos, esta última empeora en forma marcada el rendimiento cognitivo y la puntuación de síntomas totales en los pacientes con diabetes tipo 1. 17 Es más, la administración de dosis bajas de etanol retrasa la recuperación de la hipoglucemia, al reducir la producción de glucosa, demorar la respuesta de la epinefrina y predisponer a una hipoglucemia más severa. 18 Hay mayor declinación en la función cognitiva en los diabéticos tipo 1 durante los decrementos rápidos en la glucemia versus los lentos, los cuales se observan principalmente en aquellos los individuos conscientes, mientras que los inconscientes están parcialmente protegidos contra este fenómeno. 19 Cuando se compararon la insulina humana regular con la aspartato insulina administrada en forma preprandial por 16 semanas en los individuos con diabetes tipo 1, hubo incidencia global similar de hipoglucemia pero merma en los episodios nocturnos severos con la segunda. 20 Hay una asociación entre el intervalo QTc prolongado en el electrocardiograma y la hipoglucemia nocturna espontánea, debido en parte a los niveles bajos de potasio y elevados de epinefrina. 21 El período de sueńo puede predisponer especialmente a la hipoglucemia, como en individuos no diabéticos estudiados durante la hipoglucemia inducida por insulina durante el comienzo del sueńo y en el mismo momento de la noche mientras se mantuvieron despiertos, que evidenciaron un nivel más bajo de glucosa requerido para el inicio de la respuesta contrarregulatoria. Esto explicaría el riesgo aumentado de hipoglucemia severa durante el sueńo en los pacientes diabéticos. BIBLIOGRAFIA
|
|
|
|
|
|
|
|
Introduçăo Uma das principais características das células é a capacidade de manter seu meio interno com concentraçőes relativamente estáveis de íons que estăo presentes no ambiente extracelular. Esta propriedade advém da existęncia de certos tipos de transportadores protéicos (transportadores ativos primários) que, situados na membrana plasmática e utilizando-se da energia potencial contida em ligaçőes intramoleculares de alta energia, conseguem carrear espécies iônicas contra seus gradientes eletroquímicos e conservar tais distribuiçőes assimétricas. Os principais geradores deste gradiente transmembranar săo as ATPases do tipo P, responsáveis pelo transporte específico de determinados cátions, dentre eles o Na+ e o K+, de fundamental importância para a fisiologia celular. A primeira demonstraçăo da existęncia de uma enzima que hidrolisasse ATP e fosse estimulada por Na+ e K+ surgiu na metade da década de 1950 por Jens C. Skou84 em crustáceos. Na época, ainda năo era conhecido o mecanismo causador das diferenças de concentraçăo observadas para esses íons (Na+ mais concentrado fora da célula e o inverso para o K+). Skou, portanto, descreveu de forma elegante que a recém-descoberta Na+/K+-ATPase e a enigmática "bomba" de Na+ seriam a mesma entidade. Em virtude deste achado pioneiro e de sua longa trajetória na pesquisa dos mecanismos de transporte iônico, Skou foi agraciado com o Pręmio Nobel em 1997. As conseqüęncias fisiológicas desse transporte săo, direta ou indiretamente, muito relevantes para o desempenho e viabilidade celular. Assim, é possível a manutençăo do equilíbrio osmótico celular, a preservaçăo do potencial de repouso de diversos tipos celulares e das propriedades elétricas de células excitáveis. Ademais, influencia o transporte de outros íons e moléculas acoplados ao gradiente de Na+ (e.g. contratransporte de Ca2+ e H+, cotransporte de ânions como Cl-, PO43-, SO42-), substratos (e.g. cotransporte de aminoácidos neutros e glicose), e cotransporte de neurotransmissores. Outras funçőes mais restritas contudo importantes também săo mediadas pela enzima, como a remoçăo do excesso de K+ extracelular por células musculares e gliais no sentido de evitar distúrbios na excitabilidade celular.89,94 Estrutura: subunidades A subunidade a, também conhecida como subunidade catalítica ou funcional, é uma proteína integral de membrana com provavelmente 10 domínios transmembranares (politópica; massa molecular de 112 kDa) e que foi seqüenciada para vários mamíferos, como o rato80 e o homem.32 Contém os sítios de ligaçăo para cátions (Na+, K+) ATP (complexo Mg2+- ATP), fosforilaçăo e glicosídios cardíacos, uma classe de esteróides que atuam como inibidores específicos da enzima (por exemplo, a ouabaína). Esta subunidade tem propriedade ionoforética, com oclusăo e translocaçăo de íons.23 O sítio de fosforilaçăo e de ligaçăo de nucleotídios săo altamente homólogas ŕquelas presentes em outras ATPases do tipo P transportadoras de diversos cátions (Cd2+, K+, H+, Ca2+).46 Ambas as terminaçőes da proteína, amino e carboxi, estăo voltadas para a regiăo intracelular. Algumas espécies animais apresentam Na+/K+-ATPases resistentes ŕ açăo dos glicosídios cardíacos, desde invertebrados - hidra,15, anęmona-do-mar,86 camarăo marinho, 61 esquistossoma mansoni,85 borboleta monarca28 - a vertebrados como certos anfíbios do gęnero Bufo49 e determinados roedores.39,50 Esse perfil está relacionado aos aminoácidos da porçăo N-terminal, especificamente entre os segmentos transmembranares H1 e H2. Por exemplo, a mudança de dois aminoácidos de uma subunidade a sensível (carneiro) por aminoácidos carregados conferem resistęncia ŕ ouabaína.62 A presença de formas resistentes e sensíveis aos glicosídios cardíacos em organismos mais primitivos indica que a divergęncia dos genes que codificam a Na+/K+-ATPase ocorreu precocemente durante o processo de evoluçăo.90 A subunidade b, por vezes referida como subunidade regulatória, é uma proteína menor, bitópica do tipo II com maior parte localizada no meio extracelular,56 e sua massa (35-55 kDa) depende da quantidade de N- glicosilaçőes, possíveis em tręs locais diferentes. Já seqüenciada para o rato81 e o homem,31 sua homologia é extremamente alta entre mamíferos (cerca de 95%), decrescendo quando comparada com espécies de outras classes (cerca de 60%).46 Ao contrário da subunidade g, ela é essencial para a atividade Na+/K+-ATPásica, sendo co-responsável pela modulaçăo das afinidades enzimáticas ao Na+ e ao K+.9,12 A reduçăo química das ligaçőes dissulfeto impede a atividade ATPásica.47 A despeito dos esforços atualmente empregados no sentido de investigar com maior profundidade as subunidades g e, principalmente, b e suas isoformas, grande parte do conhecimento acerca da Na+/K+-ATPase está concentrada na subunidade a, por suas próprias características acima destacadas. Portanto, a subunidade a será descrita com mais detalhes a seguir, em detrimento das outras subunidades. Ademais, esta subunidade foi a única considerada neste estudo, reforçando a falta de relevância em pormenorizar as particularidades de b e g. Estrutura e localizaçăo: isoformas da subunidade a 88 Neste, foram detectadas duas formas moleculares da subunidade a através de eletroforese em gel de poliacrilamida com SDS (SDS- PAGE), que apresentavam diferença de mobilidade eletroforética µ, sendo a (forma renal - também encontrada no cérebro - e de maior µ) e a+ (forma cerebral e de menor µ), e de afinidade ŕ ouabaína (a isoforma a+ exibia afinidade 100-1000 vezes maior).88 Além disso, a isoforma a é mais sensível ŕ tripsina e mais resistente ŕ piritiamina (derivado da vitamina B) e ŕ N- etilmaleimida.40,88 Entretanto, somente em 1986, Shull et al. 80 (ver acima) foram capazes de clonar as isoformas da subunidade a de ratos, denominadas a1, a2 e a3, codificadas por genes variados. Posteriormente, tornou-se indubitável que a1 correspondia a a conferida por Sweadner, e a2 e a3 eram de fato a+. Atualmente é reconhecida a existęncia de uma quarta isoforma, a4, identificada originariamente por Shamraj e Lingrel78 e com alguns aspectos funcionais já revelados em especial pela utilizaçăo de sistemas de transfecçăo (células de inseto Sf-9 ou de mamíferos NIH 3T3), como sua alta afinidade aos glicosídios cardíacos, semelhante ŕs isoformas a2 e a3.11,97 A distribuiçăo dessas isoformas é complexa, onde por vezes, em um mesmo órgăo, células distintas comportam diferentes padrőes de expressăo. A localizaçăo subcelular também pode ser peculiar, com a expressăo de uma única isoforma, ou mais isoformas, em locais delimitados da membrana plasmática. A isoforma a1 é encontrada em todas as células de mamíferos, sendo chamada de "dona de casa", enquanto que as isoformas a2, a3 e a4 possuem uma distribuiçăo mais limitada,10,78,82 e por isso acredita-se que essas últimas sejam as responsáveis pela adaptaçăo celular permitindo ŕ célula modificaçőes no grau de expressăo dependo das exigęncias do meio. A isoforma a2 é expressa em maior abundância no miocárdio, cérebro (glia), panículo adiposo e músculo esquelético e liso,51,52,78,82 sendo igualmente detectada em cartilagem.48 A isoforma a3 é exclusiva de neurônios e outros tecidos nervosos como glândula pineal e retina,14,43,44,83 e no coraçăo do rato neonato, no qual seu nível decai ao longo do desenvolvimento, localizando-se preferencialmente no tecido condutor. 2,37,98 (Esta localizaçăo parece depender năo somente da espécie como talvez também da raça do animal, já que no ser humano95 e no rato Wistar Kyoto (WKY)65 uma maior proporçăo de isoforma a3 foi descrita o que poderia indicar uma localizaçăo muscular desta isoforma.) Entretanto, pequenas quantidades foram encontradas em cartilagem48 e endotélio.42. Nos testículos está situada a4, mais especificamente na parte medial da cauda do espermatozóide.97 A despeito de năo ter sido bem aceita quando na sua descoberta, a versăo truncada da isoforma a1 (a1 T), obtida através de variaçőes do processo de transcriçăo, parece ser uma forma constitucional do músculo liso vascular do căo45 e no ducto deferente de cobaio.27 Sua expressăo, até entăo, figura-se espécie- e órgăo-específica. Uma das dificuldades de admissăo desta isoforma é que o corte alternativo do ARNm o qual é submetida resulta em uma proteína quase 50% menor que a original (66 kDa), sendo idęntica a a1 até o resíduo Gly554 seguida por 27 aminoácidos que formam a regiăo C-terminal, enquanto que a1 full-length estende-se por mais 469 aminoácidos. Ainda assim, suas propriedades básicas como atividade ATPásica e transporte iônico foram evidenciadas.1 Importância fisiológica e regulaçăo A baixa afinidade aparente ao Na+ e ao K+ da isoforma a3, por exemplo, parece ser uma adaptaçăo evolutiva ŕ demanda fisiológica de células excitáveis. Em neurônios, ela funcionaria de forma significativa após a despolarizaçăo e gęnese de potenciais de açăo, quando os gradientes de concentraçăo estariam alterados, enquanto a1 e/ou a2 estariam atuando em níveis máximos. Portanto, a3 agiria como "bomba de reserva" na restauraçăo do potencial de repouso. Sua alta afinidade pelo ATP, outrossim, contribuiria para utilizaçăo de pequenas quantidades do nucleotídio, que estaria sendo consumido por intensa atividade neuronal. Esse mesmo raciocínio parece válido para condrócitos, que recuperar-se-iam de flutuaçőes dos níveis iônicos (mormente Na+) causadas por açăo mecânica em articulaçőes.92 Por outro lado, a2 estaria associada a captaçăo de K+ do meio extracelular. Após séries de potenciais de açăo, o aumento da concentraçăo deste íon no líquido intersticial seria anulado pela rápida interceptaçăo de a2 de astrócitos e oligodendrócitos, impedindo uma eventual despolarizaçăo indesejada. A importância fisiológica das isoformas pode também ser considerada a partir da forma como elas săo ajustadas mediante a condiçőes fisiológicas ou patológicas diversas. Ontogęnese Nos rins há duplicaçăo do número de Na+/K+-ATPases, isoforma a1, predominantemente até a terceira semana pós- natal, com estreita correlaçăo entre os valores de atividade enzimática, quantidade de a1 imunorreativa e de ARNm.36,58 No sistema nervoso central o período de incremento da densidade Na+/K+-ATPásica é semelhante. Entretanto, o aumento é manifestamente maior - tanto a atividade Na+/K+-ATPásica74 quanto a ligaçăo ŕ [3H]ouabaína73 aumenta cerca de 10 vezes durante essa fase, assim como os níveis protéicos74 e de ARNm58 das isoenzimas. A isoforma a3 é a mais abundante no cérebro do feto, enquanto que no adulto as tręs isoformas săo quantitativamente equivalentes.93 Estruturas encefálicas distintas parecem contribuir diferentemente para o aumento da quantidade de isoformas. Assim, por exemplo, isoformas de alta afinidade ŕ ouabaína apresentam maior taxa de aumento no tronco cerebral comparado aos hemisférios cerebrais, em contraste com o que ocorre para a1. 36 No músculo esquelético o ARNm de a1 é predominante no feto, mantendo níveis constantes até a idade adulta. Por outro lado, o ARNm de a2, minoritário no feto, aumenta cerca de 10 vezes transformando a2 na isoforma predominante no adulto.58 Um dos casos mais interessantes acontece no ventrículo de ratos. A isoforma a1 é a de maior expressăo no coraçăo em todas as faixas etárias do rato. Há, entretanto, uma mudança no perfil de expressăo de a2 e a3 cardíacos. No feto e no neonato a expressăo da isoforma a3 é maior que a de a2, sendo posteriormente trocado este padrăo no tecido adulto.1,37 O mesmo padrăo é verificado para o ARNm.58 A isoforma a3, em adultos, somente é encontrada no tecido condutor cardíaco. No adulto, a expressăo de isoenzimas de alta afinidade (a2 + a3) é quase 4 vezes menor que na etapa perinatal.1,36 Agentes endógenos A aldosterona, corticóide secretado pelo córtex adrenal, foi o primeiro hormônio a ser estudado por sua indiscutível açăo sobre a Na+/K+-ATPasease localizada nas células da regiăo distal do néfron, apesar de suas açőes corresponderem também a outros órgăo. A atividade Na+/K+-ATPásica de células epiteliais renais tratadas in vitro com esse mineralocorticóide sofre rápida estimulaçăo, normalmente pelo aumento do número de Na+/K+-ATPases ativas * seja pela inserçăo de enzimas pré-formadas na membrana celular ou ativaçăo de inativas já presentes.21,77 O aumento tardio da atividade Na+/K+- ATPásica está relacionado a conhecida interaçăo do esteróide com seu receptor intracelular, migraçăo ao núcleo celular e a estimulaçăo da transcriçăo genética pelo complexo formado.29 Em geral, a transcriçăo estimulada é a da isoforma a1, contudo, no hipocampo de rato foi revelada exclusivamente a elevaçăo da quantidade de ARNm da isoforma a3.19 Além de seus efeitos notórios sobre a homeostase de carboidratos, a insulina coopera para a diminuiçăo das eventualmente elevadas concentraçőes de K+ pós-prandiais, agindo sobre musculatura esquelética, fígado e rins.8 A relaçăo direta entre esses mecanismos e a Na+/K+-ATPase é indubitável. A estimulaçăo de trocadores (e.g., Na+/H+ e Na+- K+/2Cl--) e canais iônicos (canais de Na+ dependentes de insulina) confere uma rápida elevaçăo da atividade Na+/K+-ATPásica devido ŕ elevaçăo da concentraçăo de Na+ intracelular.69,72 Em células do túbulo contornado proximal renal de rato, que expressam basicamente a isoforma a1, também observou-se aumento da atividade Na+/K+-ATPásica pelo simples aumento da afinidade aparente ao Na+.20 No músculo esquelético de anfíbios e mamíferos, por outro lado, há um recrutamento de enzimas de compartimentos intracelulares.30,57. O efeito tardio na atividade e expressăo das isoformas da Na+/K+-ATPase foi verificado no músculo liso vascular, no qual há aumento da atividade Na+/K+-ATPásica e da fraçăo de ARNm da isoforma a2 91 e em astrócitos, onde há induçăo seletiva tanto da atividade quanto da traduçăo da proteína referente a a1.41 O hormônio L-triiodotironina (T3) atua de forma órgăo-específica na estimulaçăo da expressăo das isoformas da Na+/K+-ATPase, agindo năo somente sobre a transcriçăo, mas também sobre o grau de estabilidade do ARNm e de seu transporte até os ribossomos. Assim, no músculo esquelético há induçăo das isoformas a2 e b2, no rim de a1 e b1, e no coraçăo de a2 (preferencialmente), a1 e b1 - adulto - e a2, a3 e b1 - neonato.6,22,25,59,96 O sistema nervoso central de animais adultos é insensível ao hormônio, mas durante o período perinatal T33 influencia tanto enzimas de baixa quanto alta afinidade aos glicosídios cardíacos.75 Outros agentes endógenos săo igualmente importantes na organizaçăo de padrőes de expressăo adaptativos. A dopamina reduz a reabsorçăo tubular de Na+ no túbulo contornado proximal, ramo ascendente da alça de Henle cortical e tubo coletor, através, em parte, da inibiçăo da Na+/K+-ATPase basolateral.7 Esta operaçăo é mediada por uma fosfoproteína dependente de dopamina e AMPc, ou DARPP-32, que é um inibidor endógeno da proteína fosfatase 1 (PP1). A desfosforilaçăo (e ativaçăo) da Na+/K+-ATPase pela PP1 é portanto inviabilizada, permanecendo por mais tempo sob a forma inativa (fosforilada).3 A adrenalina, noradrenalina, vasopressina (ADH) e angiotensina II, ao contrário da dopamina, estimulam a atividade Na+/K+-ATPásica hepática e renal por açăo inversa, ou seja, ativaçăo indireta de fosfatase dependente de Ca2+-calmodulina (calcineurina).4,38 Fármacos colinomiméticos (e.g. carbacol) promovem aumento da atividade Na+/K+-ATPásica em células lacrimais acinares pela translocaçăo de isoenzimas a1b1 de compartimentos intracelulares para a membrana plasmática.34 Condiçőes patológicas Consideraçőes finais
|
||
|
|
| |
|
|
|
Introducción En la década de los '90 se hizo patente la importancia del bloqueo del sistema renina- angiotensina-aldosterona en la prevención de complicaciones cardiovasculares, especialmente en la hipertensión arterial y la insuficiencia cardíaca congestiva. En este último caso, la introducción de los inhibidores de la enzima de conversión de angiotensina (IECA) lograron reducir no sólo la morbilidad sino también la mortalidad de los pacientes.1 El cor pulmonale crónico (CPC) se caracteriza por una combinación de hipertrofia y dilatación del ventrículo derecho (VD), secundarias a hipertensión pulmonar (HTP) ocasionada por trastornos parenquimatosos o circulatorios pulmonares de larga evolución y etiología variada.2 El hecho fisiopatológico común a todas las formas de CPC es la hipoxia, que induce vasoconstricción e hipertensión pulmonares y finalmente hipertrofia e insuficiencia ventricular derechas.2,3 La angiotensina II, por su acción vasoconstrictora y promotora del crecimiento y división celular, es un elemento clave en el desarrollo y mantenimiento de la hipertrofia y fibrosis cardíaca.4Además, la hipoxia crónica provoca un incremento de la actividad de la enzima de conversión de la angiotensina (ECA) sérica.5 La administración de captopril solo6 o asociado a oxigenoterapia7 es capaz de reducir la presión arterial pulmonar en enfermos con EPOC así como la hipertrofia vascular y del ventrículo derecho en ratas hipóxicas.8 A pesar de la importancia del sistema RAA en la fisiopatología vascular y de algunas similitudes entre el fallo cardíaco del ventrículo izquierdo (VI) y el derecho (VD), no existen estudios en el ámbito clínico, a medio o largo plazo, que analicen el papel de los IECA en el CPC. Con ese ánimo se diseńó y realizó el estudio al cual se refiere el presente artículo.9 Diseńo del estudio Se incluyeron 28 pacientes con cor pulmonale crónico. El diagnóstico de CPC se formuló en todos los casos sobre la base de criterios clínicos o ecocardiográficos de fallo o hipertrofia del ventrículo derecho (VD). Los criterios concretos de inclusión/exclusión pueden consultarse en la publicación original.8 Todos los pacientes incluidos dieron su consentimiento por escrito, tras ser informados. El protocolo fue aprobado por el comité de ensayos clínicos del Hospital Clínico Universitario. Se administró a cada paciente enalapril o placebo según un protocolo de aleatorización. Las dosis del fármaco a estudio se suministraron comenzando a dosis bajas (2.5 mg de sustancia activa o el placebo equivalente), con aumento gradual cada semana hasta alcanzar la dosis objetivo de 20 mg, si no se presentaban problemas de tolerancia. Se dejó libremente a criterio médico el resto de la medicación. Se realizó seguimiento a 1, 3 y 6 meses de alcanzada la dosis máxima de fármaco mediante examen analítico (hematimetría y bioquímica completas) de sangre y orina, radiografía de tórax y ECG. Previamente a la administración del fármaco, y al finalizar las 24 semanas de seguimiento, se efectuó en cada paciente ecocardiografía transtorácica bidimensional, ventriculografía isotópica basal y estimulada con dobutamina, estudio de función respiratoria (FEV1, FVC, FEV1/FVC y gasometría arterial respirando aire ambiente) y valoración de la capacidad de esfuerzo mediante el test de 6 minutos caminando. Estudio estadístico Resultados
Clase funcional. De los pacientes que finalizaron el estudio, 20 estaban en clase funcional II y 6 en clase III (NYHA), en situación basal. La distribución de pacientes fue uniforme en ambos grupos. La dosis de fármaco administrado fue de 16.15 mg en el grupo de enalapril y de 20 mg en el placebo (tabla 1). Efectos adversos. Ningún paciente tuvo que abandonar el tratamiento con el fármaco (enalapril o placebo) a consecuencia de efectos adversos graves. El efecto más frecuentemente observado fue la disminución de las cifras de TA en el grupo tratado, aunque fue bien tolerado y ningún paciente precisó reajuste en las dosis. No se observaron casos de tos, ni reacciones cutáneas o edema angioneurótico en el grupo de tratamiento activo. No hubo cambios en la exploración funcional respiratoria (tabla 2).
Los cambios ecocardiográficos se reflejan en la tabla 3. Muestran una tendencia a la disminución del espesor de la pared del VD y un incremento de la fracción de eyección del VD (FEVD). Cambios en la ventriculografía isotópica basal. La FEVD al inicio en el grupo placebo fue de 41.8% y de 41.7% en el grupo enalapril. Al final del período de tratamiento fue de 39% y 44.43%, respectivamente (figura 1).
Figura 1. Variación de la FEVD tras 6 meses de tratamiento. La fracción de eyección del VI (FEVI) del grupo placebo al inicio fue de 57.38%, mientras que la del grupo enalapril fue de 58.44%. Al finalizar el estudio fueron respectivamente del 57.2% y 61.60% (tabla 3).
Al final del estudio, la FEVD disminuyó en el grupo placebo un 7.09%, mientras que en el grupo enalapril se produjo un incremento del 6.4%. La diferencia fue del 13.5% a favor del grupo enalapril (figura 2).
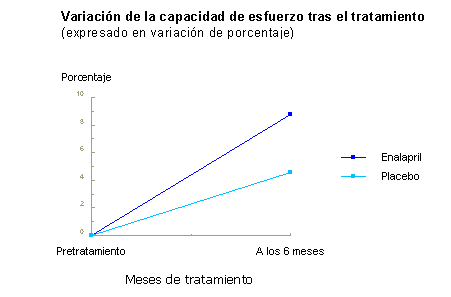
Figura 2. Variación de la FEVD, expresada en porcentaje, al final del tratamiento. Ninguna de las diferencias alcanzó significación estadística. Ventriculografía isotópica con dobutamina. Al finalizar el estudio se observó que todos excepto 2 de los pacientes del grupo placebo estaban en fibrilación auricular al final del período de seguimiento de 6 meses, mientras que tan sólo 2 del grupo de tratamiento activo presentaban esta arritmia. Puesto que esta arritmia era contraindicación para la administración de dobutamina, no pudieron obtenerse datos comparativos. Cambios en el test de "seis minutos caminando" (tabla 2 y figura 3). Demuestran una mejor tolerancia al esfuerzo en ambos grupos, con tendencia a mejorar más aquellos del grupo enalapril. Las diferencias, sin embargo no alcanzaron significación estadística.
Figura 3. Variación de la capacidad de esfuerzo, expresada en porcentaje, al final del tratamiento. Comentario Lo interesante de estos resultados es que mientras que hay una mejoría global de la FEVD (+6.4%) en el grupo enalapril, en el grupo placebo no sólo no mejora sino que se produce un deterioro al final del estudio de la misma magnitud (-7.09%). Esta diferencia se acompańó además de un incremento en la capacidad de ejercicio de los pacientes tratados, que duplicó la de los sujetos del grupo placebo. Si observamos los datos ecocardiográficos, apreciaremos que esta misma "congruencia" se mantiene. En el grupo enalapril hay disminución (aunque no significativa) del espesor de la pared y del diámetro telediastólico del VD que no se da en el grupo placebo. Sin embargo no hay cambios en la PSAP. Por otra parte, es importante reseńar la ausencia de efectos secundarios debidos al tratamiento en pacientes hipotéticamente expuestos a la hipotensión y el síncope. Llama la atención que no se registró entre los pacientes con tratamiento activo un aumento en la incidencia de tos. Aunque las diferencias expuestas no llegan a alcanzar significación estadística, marcan una tendencia congruente hacia la mejoría en el grupo de pacientes tratados con enalapril, expresada por incremento de la capacidad de esfuerzo y sustentada en una mejoría del rendimiento del VD. La ausencia de significación estadística de los resultados puede obedecer al tamańo muestral, al período de seguimiento, breve para una enfermedad crónica, y al hecho de no haber incluido pacientes con oxigenoterapia domiciliaria para evitar dispersión de los resultados. En conjunto, la congruencia de los resultados indica que el enalapril asociado al tratamiento convencional del CPC podría contribuir en la prevención el deterioro progresivo de la función sistólica del VD. Estos efectos parecen ejercerse primariamente sobre el músculo cardíaco, ya que no se acompańa de modificaciones en la función respiratoria, gasometría, función renal ni hemodinámica pulmonar. Siglas: IECA: inhibidores de la enzima de conversión de angiotensina. CPC: cor pulmonale crónico. VD: ventrículo derecho. HTTP: hipertensión pulmonar. VI: ventrículo izquierdo. VD: ventrículo derecho. FEVD:fracción de eyección del VD. PAP: presión arterial pulmonar. PSAP: presión sistólica arterial pulmonar. BIBLIOGRAFIA
|
Trabajos Distinguidos, Serie Clínica
Médica,
integra el Programa SIIC de Educación Médica Continuada
![]()
Bienvenidos
a siicsalud
[Acerca
de SIIC] [Estructura de SIIC]
[Círculo de Lectores
SIIC]
[Salud(i)Ciencia]
[Trabajos Distinguidos]
Sociedad
Iberoamericana de Información Científica (SIIC)
Av. Belgrano 430, (C1092AAR), Buenos Aires, Argentina
Correo electrónico (e-mail): atencionallector@siicsalud.com;
Tels: +54 11 4342-4901; Fax: +54 11 4331-3305.
Correo SIIC: Casilla de Correo 2568, (C1000WAZ) Correo Central, Buenos Aires.
Copyright siicsalud©
1997-2003, Sociedad Iberoamericana de Información Científica (SIIC)