Volumen 18, Número 3, Junio 2004

Federación
Argentina de Sociedades de Ginecología
y Obstetricia (FASGO)
![]()
|
|
|
|
|
|
||
|
Expertos Invitados |
|
|
|
|
 |
Columnista
Experto de SIIC Dr. Richard M. Grimes Associate Professor of Management and Policy Sciences, School of Public Health at the University of Texas Houston Health Science Center, Houston, Texas, EE.UU.Participaron en la investigación: Nancy L. Eriksen MD y Andrew W. Helfgott. |
Introducción Se acepta que el estudio serológico para la detección de infección por HIV durante el embarazo puede asociarse con beneficios potenciales, tanto para la madre como para el feto. Entre las ventajas hay que mencionar la posibilidad de iniciar el tratamiento en forma precoz, lo cual es de gran beneficio. De hecho, existe evidencia sustancial de que el tratamiento apropiado precoz de la infección por HIV mejora la evolución de los pacientes con terapia antirretroviral. Por lo tanto, la detección de la infección por HIV en la gestación permite que la madre pueda ser tratada más tempranamente y que reciba el tratamiento antiHIV que reduce la morbilidad y aumenta la sobrevida.1 Además, la identificación prematura de la infección también permite que la mujer reciba la terapia profiláctica para infecciones oportunistas asociadas con la infección por HIV. Está establecido que la medida prolonga y mejora la calidad de la vida.2 Conocer el estado de infección por HIV de una mujer embarazada también tiene un beneficio para el nińo. La transmisión del HIV de madre a hijo es aproximadamente del 40% en las mujeres que amamantan. Se considera que la eliminación de esta forma de alimentación reduce el índice en alrededor de un 14%.3 También es sabido que es mucho menos probable que las mujeres tratadas con antirretrovirales durante la gestación transmitan el virus a la descendencia. El estudio clásico (llamado protocolo 076) mostró que las madres e hijos que reciben monoterapia con zidovudina (AZT) tienen reducción del riesgo de transmisión del orden del 25.6% al 8.3%. Las mujeres del estudio recibieron AZT oral antes de la semana 34 de gestación y en forma intravenosa durante el trabajo del parto. Asimismo, los hijos recibieron AZT en jarabe desde inmediatamente después del nacimiento y durante las seis semanas posteriores.4 También hay evidencia de que la práctica obstétrica puede reducir el riesgo de transmisión materno-fetal del HIV. Landesman y colaboradores mostraron que la ruptura prematura de membranas se asocia con mayor probabilidad de transmisión del HIV.5 Además, un metaanálisis del riesgo de transmisión mostró que el uso de AZT junto con la cesárea programada y la no alimentación a pecho pueden reducir el riesgo de transmisión a un escaso 2%.6 Debido a que el conocimiento del estado de infección por HIV en mujeres gestantes tiene enormes consecuencias para la madre y para el nińo, muchos programas gubernamentales recomiendan el estudio serológico como parte de la atención prenatal. El estado de Texas, en los Estados Unidos, promulgó una ley para que el profesional ofrezca a la mujer la posibilidad de estudio serológico en la primera visita y en el momento del parto. La conducta se asocia con el beneficio de poder comenzar con la terapia antirretroviral antes del nacimiento, de evitar la ruptura prematura de membranas y de planificar la cesárea, con la finalidad de reducir el riesgo de transmisión materno-fetal. Asimismo, brinda la posibilidad de comenzar el tratamiento antiviral en la madre lo antes posible y permite al profesional aconsejar sobre las formas más seguras de alimentación del recién nacido. El requisito de repetir el estudio serológico en el momento del parto asume la posibilidad de que un determinado número de mujeres seroconvierte durante la gestación. Nuevamente, la información en este momento es útil para desalentar la lactancia materna. Primera pesquisa de infección materna Sin embargo, el índice de seroconversión durante el embarazo no era conocido. Por ende, los autores realizaron un estudio en 97 mujeres que tuvieron serología positiva en el momento del parto, durante 1996 y 1997, en dos hospitales académicos del University of Texas Houston Health Science Center. Se revisaron las historias clínicas de las mujeres para saber si habían tenido un resultado positivo en la etapa de atención prenatal, con lo cual podrían determinar la seroconversión durante la gestación. No encontraron resultados compatibles con seroconversión durante el embarazo; 30 de las 97 mujeres sabían que estaban infectadas antes de la concepción por lo que el primer estudio y la repetición en el momento del parto no proporcionaron información útil. La ausencia de seroconversión sugirió, además, que la reevaluación posterior sería de poco valor. No obstante, 21 de las 67 mujeres que desconocían su estado de infección antes del embarazo no recibieron atención prenatal (n = 11) o recibieron el primer control después de la semana 34 de gestación (n = 10). Esta edad gestacional es importante porque el protocolo 076 acerca del uso de AZT en embarazo fue realizado en pacientes que recibieron el antiviral antes de la semana 34 de gestación. Por lo tanto, para ese entonces se consideraba crucial la valoración antes de esa fecha de gestación. Sobre esa base, en 10 de las 67 (15%) mujeres con diagnóstico reciente, la detección de la infección se realizó demasiado tarde como para recibir el beneficio máximo de la terapia antirretroviral. Sin embargo, el diagnóstico en ese momento aun permitió evitar el riesgo relacionado con la ruptura prematura de membranas y con la lactancia y programar la cesárea. En un 16% adicional (n = 11) de la cohorte, el diagnóstico de infección por HIV se efectuó demasiado tarde como para que las medidas terapéuticas redujeran el riesgo de infección fetal a partir del tratamiento antiviral, la ruptura prematura de membranas y la cesárea electiva. Sin embargo, todavía hubo tiempo de evitar la lactancia. Así, la detección del estado de infección por HIV al inicio del cuidado prenatal y nuevamente en el momento del parto tiene algún beneficio en relación con la reducción potencial de transmisión perinatal del HIV. El estudio reveló la importancia de efectuar el estudio serológico en el momento del parto en mujeres que no habían recibido cuidados prenatales. Además, quedó demostrada la importancia de aconsejar y educar a las mujeres que tienen riesgo de infección por HIV a que soliciten atención prenatal en forma precoz. Los esfuerzos deberían formar parte de cualquier programa integral destinado a reducir la transmisión del HIV de madre a hijo durante el período perinatal.7 Estudios de seguimiento Desde la terminación del estudio en 1997, varias investigaciones demostraron el valor de conocer el estado de infección por HIV en la madre, aun en forma tardía durante la gestación. Un trabajo realizado en Tailandia reveló que el índice de transmisión viral de la madre al feto era del 8.3% cuando la madre HIV positiva iniciaba el tratamiento con AZT en la semana 35 de la gestación y cuando los hijos recibían la droga durante tres días después del nacimiento.8 Los hallazgos se confirmaron en un estudio que comparó mujeres que iniciaron el tratamiento antiviral entre las semanas 36 a 38 de la gestación y aquellas que recibieron placebo. El índice de transmisión en este último grupo fue del 27.5% versus un 18% en el grupo activo.9 Un trabajo similar, de Uganda, mostró que la administración de nevirapina o de AZT a la madre, durante el trabajo de parto, y a sus hijos después del nacimiento, podía reducir el índice de transmisión del HIV a un 8.2% (en el grupo tratado con nevirapina) y a un 10.4% en el grupo que recibió AZT. En el primer grupo (nevirapina), las madres recibieron una única dosis oral al inicio del trabajo de parto y los hijos recibieron una única dosis dentro de los tres primeros días de vida. Las madres tratadas con AZT recibieron una combinación oral e intravenosa del medicamento y los infantes fueron tratados con dos dosis diarias durante 7 días después del nacimiento.10 Wade y colaboradores realizaron un estudio retrospectivo acerca del efecto de la administración de AZT a la madre en el momento del parto, y al nińo después del nacimiento, sobre la transmisión de la infección. Encontraron que el protocolo 076 completo no era necesario para lograr una reducción sustancial del índice de transmisión de madre a hijo. Cuando las mujeres recibieron el protocolo 076 completo, el índice de contagio fue del 6.3%. Sin embargo, si la madre sólo recibía AZT durante el parto, el índice de transmisión era del 10%. En caso de que el nińo comenzara a recibir AZT dentro de las 48 horas posteriores al nacimiento, el índice de infección era del 9.3% mientras que se elevaba a un 18.4% cuando el tratamiento se iniciaba al tercer día del parto o en forma más tardía. En ausencia de profilaxis con AZT, el índice de transmisión fue del 26.6%.11 Todos los trabajos mostraron la importancia de descubrir la infección en la madre durante el embarazo avanzado. Además, revelaron la importancia de reconsiderar el valor de determinar la seroconversión durante la gestación. Los profesionales del Centro Inmunológico de Mujeres (NLE), en uno de los dos hospitales del estudio original, continuaron controlando la población de mujeres embarazadas asistidas en la clínica con el objetivo de detectar seroconversión antes del estudio prenatal y en el momento del parto. Durante enero de 1997 y febrero de 2002, 180 mujeres de las cuales pudieron revisarse las historias clínicas fueron positivas en el momento del parto. Entre ellas, tres que habían sido seronegativas en el primer control prenatal fueron positivas en esa instancia. El seguimiento confirmó el beneficio de repetir la serología, que no se había constatado en el estudio original. La posibilidad de seroconversión durante el período de atención prenatal quedó confirmada. Recomendaciones para la obtención de máximos beneficios El rastreo de infección por HIV en el momento del parto duplica los costos de la determinación perinatal. Durante el parto, la determinación representa un gasto de apenas 16 dólares, incluidos los honorarios del técnico.12 Debido a que en otros países el costo del parto es aun menor, también es posible que el gasto total sea incluso más bajo. Si bien no es un gasto trivial, el segundo estudio serológico parece asociarse con un beneficio real para un número pequeńo de mujeres y sus hijos. El rastreo en el momento del nacimiento parece de valor particular para nińos de madres que no reciben o que reciben escasa atención prenatal. Además, el estudio de seguimiento sugiere que puede existir cierto beneficio del rastreo en el momento del parto porque existe la posibilidad pequeńa pero real de seroconversión durante el período de atención prenatal. De esta manera, al obviar un segundo estudio, un número de nińos nacidos de madres HIV positivas están en riesgo de contagio del virus a través de la alimentación a pecho. Además, en caso de que la seropositividad de la madre no se reconozca en el momento del parto, se anula la posibilidad de falta de infección del recién nacido a partir del tratamiento de la madre y del nińo con AZT. El beneficio de conocer el estado de infección de la madre en el momento del parto también es ventajoso para ella, ya que puede comenzar a recibir terapia antirretroviral y profilaxis para las infecciones oportunistas en forma más temprana. La información puede ser útil también para asesorar a las mujeres en relación con la prevención de futuros embarazos o con la posibilidad de recibir cuidados prenatales tempranos en caso de una nueva gestación. Sin embargo, para lograr el máximo beneficio del estudio serológico en el momento del nacimiento, debe estar garantizada la posibilidad de implementar todas las medidas correspondientes que surgen de la identificación del estado positivo, entre ellas la disponibilidad de AZT para ser administrado a la madre y al hijo. El sistema de rastreo debe estar disponible y el resultado debe ser rápidamente comunicado al profesional, de manera tal que el tratamiento con AZT pueda iniciarse con la mujer aun en trabajo de parto, con lo cual todavía hay tiempo de programar la cesárea y evitar la ruptura prematura de membranas. Aun cuando el laboratorio no esté en condiciones de dar los resultados en forma tan rápida, disponer de ellos es ventajoso porque permite adoptar las medidas correspondientes para iniciar el tratamiento en el nińo antes de las 48 horas posteriores al parto. Sin embargo, ningún procedimiento es de utilidad si no hay AZT disponible para administración intravenosa o en forma de jarabe. Por lo tanto, el rastreo serológico debe formar parte de un sistema organizado de cooperación entre el laboratorio, la farmacia y el equipo de obstetricia. También hay que mencionar que en una proporción significativa de mujeres en quienes el estado de infección por HIV se descubre en el momento del parto (aquellas con escasa o nula atención prenatal), el diagnóstico es demasiado tardío como para obtener los máximos beneficios para el nińo. El diagnóstico pudo haberse efectuado demasiado tarde como para obtener los beneficios de la terapia antirretroviral, para evitar la ruptura prematura de membranas y para programar la cesárea. Para la mayoría de estas mujeres, la evitación de la lactancia y el tratamiento del recién nacido son las únicas intervenciones posibles destinadas a reducir el riesgo de transmisión materno-fetal. Esto pone de manifiesto la enorme importancia de la educación de todas las mujeres embarazadas en relación con el valor del control prenatal temprano, especialmente para aquellas en riesgo de infección por el HIV. Asimismo, pone de relieve la necesidad de que las clínicas prenatales brinden la mayor posibilidad de atención, en relación con las horas de trabajo, el personal disponible y la localización. Cada mujer que no tiene posibilidad de acceso a un servicio de atención prenatal, ya sea por ignorancia o porque éste no está a su alcance, representa una fuente de contagio del HIV a su descendencia. Proporcionar la posibilidad de rastreo y tratamiento no es suficiente: los centros específicos deben asegurar el medio prenatal adecuado para que puedan obtenerse los máximos beneficios posibles.
2. 2001 USPHS/IDSA Guidelines for the prevention of opportunistic infection in persons infected with the human immunodeficiency virus. Found at http://hivatis.org/guidelines. 3. Dunn DT, Newell ML, Ades AE, Peckham CS. Risk of human immunodeficiency virus type 1 transmission through breastfeeding. Lancet. 1992;340:585-88. 4. Connor EM, Sperling RS, Gelber R, et al. Reduction of maternal-infant transmission of human immunodeficiency virus type I with zidovudine treatment. N Engl J Med. 1994;331:1173- 80. 5. Landesman SH, Kalish LA, Burns DN, C et al. Obstetrical factors and the transmission of Human immunodeficiency virus type 1 from mother to child. N Engl J Med 1996;334:1617-23. 6. Perinatal HIV Guidelines Working Group. Recommendations for Use of Antiretroviral Drugs in Pregnant HIV-1 Infected Women for Maternal Health and Interventions to Reduce Perinatal HIV-1 Transmission in the United States. Found at www.hivatis.org. 7. Grimes RM, Srivastava G, Helfgott AW, Eriksen NL. A study to determine the incidence and prevalence of newly discovered Human immunodeficiency virus infection during the prenatal care period. Infect Dis Obstet Gynecol. 2000;8:172-5. 8. Lallemant M, Jourdain G, Le Cour S et al. A trial of shortened zidovudine regimens to prevent mother-to-child transmission of human immunodeficiency virus type 1. N Engl J Med 2000;343:982-91. 9. Dabis F, Msellati P, Meda et al. 6-month efficacy, tolerance and acceptability of a a short regimen of oral zidovudine to reduce vertical transmission of HIV in breastfed children in a Cote d'Ivoire and Burikino Faso: a double blind placebo-controlled multi-centre trial. Lancet. 1999;353:786-92. 10. Guay LA, Musoke P, fleming T et al. Intrapartum and neonatal single dose nevirapine compared with zidovudine for preventionof mother-to-child transmission of HIV-1 in Kampala Uganda. Lancet. 1999;354:795-802. 11. Wade NA, Birkhead GS, Warren BL et al. Abbreviated regimens of zidovudine prophylaxis and perinatal transmission of the human immunodeficiency virus. N Engl J Med. 1998;339:1409-14. 12. Phillips KA, Lowe RA, Kahn JG et al. The cost effectiveness of HIV testing physicians and
|
|
|
|
|
 |
Columnista
Experto de SIIC Augusto Simőes-Barbosa Professor/pesquisador em Patologia molecular. Programa de Pós-graduaçăo em Cięncias Genômicas e Biotecnología, Universidade Católica de Brasilia, Brasil |
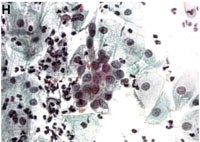
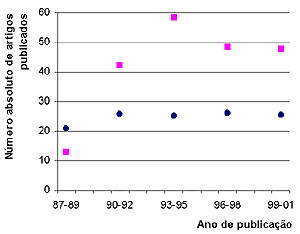
Doenças sexualmente transmissíveis, um problema mundial a ser enfrentado Doenças sexualmente transmissíveis (DST) săo causas importantes de morbidade e mortalidade em todo o mundo, e vem experimento aumento no registro de casos a partir da década de 1950 devido a um complexo de fatores da vida moderna. De maneira ilustrativa, ocorrem anualmente em todo o mundo 170 milhőes de casos de infecçőes por Trichomonas vaginalis, 89 milhőes por Chlamydia trachomatis, 62 milhőes de casos de gonorréia, 12 milhőes de sífilis, 30 milhőes de condilomas anogenitais, 20 milhőes de herpes anogenital e 2 milhőes de cancróide. Atualmente, para a AIDS, estimam-se 30.6 milhőes de casos.1 Por exemplo, as infecçőes por Chlamydia e a sífilis săo consideradas grandes fontes de morbidade e mortalidade, estando entre as quinze primeiras causas de incapacidade ou dano ŕ saúde humana em populaçőes urbanas.2 Outro exemplo advém do câncer cervical, que tem sua etiopatogęnese determinada pela infecçăo de subtipos carcinogęnicos de HPV, e incidęncia de 500 000 casos/ano.3 Em média, a incidęncia de DST é sempre superior em pessoas entre a faixa etária de 15 a 35 anos, correspondendo em média ao período de maior atividade sexual, pertencentes a ambientes urbanos ou peri-urbanos. Nos países industrializados registra-se por ano um novo caso de DST em cada 100 pessoas, e nos países em desenvolvimento as DST estăo entre as 5 principais causas de procura por serviços de saúde.1 De fato, pobreza social e econômica săo também adjuvantes de DST no contexto de países em desenvolvimento.4 Por exemplo, 80% da incidęncia e mortalidade do câncer uterino ocorre em países em desenvolvimento.3 Sem dúvida, a grave importância de DST na saúde publica foi acelerada pela epidemia da AIDS em todo o mundo. A literatura registra que o risco de infecçăo por HIV, vírus da imunodeficięncia humana, aumenta de duas a cinco vezes sempre que as pessoas săo portadoras de DST ulcerativas, năo-ulcerativas e inflamatórias. Mesmo no último caso, micro-ulceraçőes, friabilidade da mucosa vaginal e um excesso de macrófagos e linfócitos (células hospedeiras do HIV), decorrentes da forte leucorréia, tornam-se adjuvantes para transmissăo ou aquisiçăo do vírus. Ainda, é sugerido que a diminuiçăo da flora lactobacilar produtora de peróxido de hidrogęnio permite maior sobrevivęncia do vírus no ambiente.5 De forma geral, associaçőes epidemiológicas tęm sido encontradas entre DST e infecçăo por HIV.6-8 Da mesma forma, o tratamento de populaçőes na tentativa de combate as DST pode levar a reduçăo no aparecimento de novos casos de infecçăo por HIV.9 Dados de nossos trabalhos prévios indicam uma situaçăo alarmante para o caso das DST em nossa populaçăo, situaçăo que parece ser comum a populaçőes de países em desenvolvimento.4-6 Em um estudo longitudinal de 6 anos, foi preocupante notar a alta prevalęncia de lesőes inflamatórias (65%), e que apenas uma pequena parcela de lesőes pré-cancerosas e cancerosas (6%) correspondiam a lesőes iniciais de categoria NICI (Neoplasia Intra-Cervical de grau I). Podemos inferir que grande parte das mulheres năo atendem corretamente a prevençăo ginecológica de câncer cervical, ou se fazem, os métodos de diagnóstico preventivo utilizados năo săo adequados a ponto de detectar as lesőes iniciais.10 Na verdade, nos países em desenvolvimento, apenas 5% das mulheres atendem corretamente aos serviços de triagem do câncer cervical, enquanto que nos países desenvolvidos esse número alcança 40%. Este é um dos fatores importantes relacionados ao fato de que, em países em desenvolvimento, 80%-85% das mulheres com câncer cervical atendem ŕ ginecologia em estágios incuráveis.11,12 Muitas devem falecer pela doença sem nunca terem sido atendidas pela clínica. Esse deve ser o quadro também para outras infecçőes e doenças sexualmente transmissíveis. No entanto, como pesquisadores da área em países em desenvolvimento, nós entendemos que năo só a falta de informaçăo é fator importante, mas também o difícil acesso, as condiçőes precárias ao atendimento de saúde, e, por último, os métodos insuficientes de diagnóstico também participam da dificuldade do controle epidemiológico e de progressăo dessas doenças nos indivíduos acometidos pelas infecçőes sexuais. A questăo do diagnóstico, tema a ser tratado em especial neste trabalho, é ponto chave para o correto tratamento, aconselhamento e manejo das infecçőes sexuais. O diagnóstico usual de doenças sexualmente transmissíveis Ainda que os sintomas e os exames clínicos simples possam indicar a etiologia da doença, o diagnóstico de DST deve ser baseado em procedimentos laboratoriais, uma vez que essas análises primárias diretamente sobre o paciente tem baixa prediçăo de acerto. Mesmo a colposcopia, considerada padrăo ouro para lesőes decorrentes de HPV, tem baixa especificidade (inferior a 50%), fato que leva a superestimaçăo da infecçăo e lesăo, e tratamento desnecessário, implicando em gasto e prejuízo psicobiológico e de saúde para o paciente. A junçăo de elementos sintomatológicos e dados de exames clínicos permitiu a geraçăo de poderosos algoritmos que podem facilitar o diagnóstico de DST. É a chamada abordagem sindrômica, que ainda assim năo é satisfatória, pois os sintomas e os dados clínicos isolados ou em conjunto podem năo permitir a distinçăo das doenças em muitos casos.13 Considerando as importantes resultantes patológicas de DST na populaçăo, o correto diagnóstico dessas doenças passa a ser crítico. Para que o atendimento ginecológico possa cuidar do adequado tratamento e prevençăo de DST, prenatal e planejamento familiar e que, por sua vez, a vigilância alcance resultado satisfatório no controle de doenças tăo importantes, um diagnóstico preciso ainda é necessário. O diagnóstico laboratorial de DST é geralmente baseado em um minucioso exame microscópico das células e secreçăo colhidas por descamaçăo exfoliativa ou abrasiva. O método citológico mais comum é denominado Papanicolaou, baseado na combinaçăo de tręs corantes que resultam em tonalidades variadas e, dessa forma, permite a visualizaçăo de detalhes do núcleo e citoplasma das células fixadas. O Papanicolaou tem grande apelo ao uso uma vez que é possível distinguir a etiologia de diversas lesőes inflamatórias e năo-inflamatórias num único exame, o que agiliza o processo de leitura e diminui os custos.14 Isso torna o procedimento adequável para rotina laboratorial em massa e triagem, e especialmente na alta rotina dos atendimentos públicos de saúde em grandes hospitais. A citologia do Papanicolaou permite a distinçăo de diferentes etiologias inflamatórias e suas alteraçőes citopatológicas, como Trichomonas vaginalis, micoses por Candida sp, viroses por Citomegalovírus (CMV), vírus da Herpes (HSV) e Papillomavirus humano (HPV), presença de bactérias pela flora lactobacilar, Gardnerella vaginalis, actinomicetos, Klebsiella granulomatis (sinon. Calymatobacterium granulomatis), Chlamydia trachomatis, Leptothrix vaginalis, e até cistos de amebas e ovos de helmintos.14 Em lesőes inflamatórias, é notável a presença de um exsudato inflamatório composto por leucócitos e macrófagos, eventualmente hemáceas, uma vez que a friabilidade da mucosa vaginal pode levar ao fácil sangramento durante a coleta. No entanto, ressaltamos que este método citológico tem grande limitaçőes e pode apresentar falhas consideráveis.15-17 Primeiro, deve-se tem em mente que pelo Papanicolaou năo é possível a detecçăo segura de diversas infecçőes bacterianas, sendo necessário utilizar de técnicas de isolamento e cultura microbiológica. Para estes casos, vale citar: estafilococos, estreptococos, Haemophylus ducrey e Neisseria gonorrhoeae. Ainda mais, é notoriamente reconhecido que a cultura também é padrăo ouro para Candida e Trichomonas vaginalis. O T. vaginalis pode ainda ser bem observado, dada a motilidade do parasita e também a experięncia do citologista observador, em um simples exame microscópico a fresco, desde que a observaçăo seja feita imediatamente após a colheita ginecológica. Embora possível, a sensibilidade e especificidade do diagnóstico de Chlamydia trachomatis săo insatisfatórios pela citologia.18 Outra dificuldade advém da impossibilidade de cultivo acelular desta bactéria para diagnóstico. Daí, é necessário recorrer a técnicas imunológicas, assim como para detecçăo do Treponema pallidum. Este último ainda pode ser facilitado clinicamente quando há aparecimento do cancro. No entanto, na infecçăo treponęmica há uma ampla janela imunológica de até 3 semanas após o desenvolvimento do cancro, quando os testes imunológicos săo pouco sensíveis. A microscopia em campo escuro é uma alternativa que necessita de equipamento especial, médicos e técnicos bem treinados, e ainda apresenta baixa sensibilidade sendo necessário no mínimo 105 organismos/ml. O teste de infectividade em coelhos é padrăo ouro para sífilis, de grande sensibilidade (até um único microorganismo), no entanto sendo necessário facilidades para manipulaçăo de animais.19 Na infecçăo por K. granulomatis, embora restrita a focos tropicais, năo há alternativa ŕ citologia senăo a detecçăo molecular.20 A Figura 1 apresenta alguns resultados citológicos pelo Papanicolaou e revela os critérios definitivos para detecçăo de alguns agentes infecciosos genitais.14 A infecçăo por Gardnerella vaginalis é caracterizada pela presença de numerosos pequenos bacilos parcial ou totalmente aderidos na superfície das células que tornam-se de coloraçăo azul clara. As células recobertas pelas bactérias assumem uma característica típica, e săo denominadas clue cells (Figura 1A). Candida albicans é um fungo patogęnico detectado em forma filamentosa ou de esporo. Filamentos marrom escuro ficam como galhos que se cruzam, enquanto que os esporos apresentam um halo claro devido a presença de uma parede celular bem delimitada. Essas características săo definitivas para diagnóstico da candidíase genital (Figura 1B). O protozoário Trichomonas vaginalis possui uma forma elíptica, medindo cerca de 10 a 20 micrômetros, tomando tonalidade acinzentada a azulada. É usualmente encontrado próximo ou aderido ŕs células epiteliais. Características definitivas devem ser a presença do núcleo periférico e traços do flagelo (Figura 1C). Porém, estas características podem tornar-se imperceptíveis.16 Por ser uma bactéria de vida intracelular obrigatória, a Chlamydia trachomatis é encontrada como inclusőes eosinófilas preferencialmente nas células da regiăo da junçăo escamocolunar e endocévice. Formam-se geralmente múltiplos vacúolos delimitados devido a formaçăo de um verdadeiro vacúolo citoplasmático bem definido (Figura 1D). As viroses săo marcadas por alteraçőes citológicas. O vírus da herpes genital (HSV) induz a megalia do núcleo, assim como a formaçăo de células multi-nucleadas, aonde os grandes núcleos se justapőem. O núcleo também toma um aspecto vítreo característico, possuindo inclusőes eosinófilas (Figura 1E). O HSV pode atingir e formar lesőes nas camadas de células malpighianas, metaplásicas e endocervicais.14,21 Células infectadas pelo Papillomavirus humano (HPV) desenvolvem atipias, muitas vezes discretas. A coloicitose é predominante. Refere-se ao alargamento do núcleo (relaçăo núcleo/citoplasma alterada) o qual torna-se envolvido por um halo citoplasmático claro e em forma côncava (Figura 1F). O colo do útero, em especial a zona de transformaçăo, é o local preferencial de infecçăo pelo HPV e de potencial oncogęnico. Ressalta-se que a citologia năo permite a distinçăo entre os subtipos de HPV. Dada a relaçăo oncogęncia da infecçăo pelo HPV, este é o ponto mais crítico em termos de diagnóstico de DST na atualidade. A infecçăo com HPV pode ser confundida com processos neoplásicos ou apenas metaplásicos.14,15,21 As anormalidades citológicas de baixo grau identificadas como ASCUS (células escamosas atípicas de significado indeterminado) e AGUS (células glandulares atípicas de significado indeterminado) săo muitas vezes mau interpretadas por limitaçőes técnicas (Figura 1G e 1H). Há enorme subjetividade na interpretaçăo destas atipias celulares, fato que se traduz na variabilidade de resultados inter-observador ou entre diferentes laboratórios.22 Esta subjetividade leva, na maioria dos casos, a uma super-estimaçăo do diagnóstico, tratamento desnecessário e estresse emocional para a paciente. Isso encarece os custos do rastreamento, pois estas mulheres serăo submetidas a exames complementares e a uma rotina mais freqüente de visita ginecológica. Os dados a seguir mostram o quăo conflitante é o diagnóstico do HPV e lesőes cervicais versus atipias celulares. É certo que 10%-15% das mulheres com ASCUS tenham na verdade lesőes de alto grau ou câncer. Também, mais da metade das lesőes de alto grau săo precedidas por diagnóstico citológico de ASCUS ou AGUS.3,23 O erro intrínseco da técnica do Papanicoloau é da ordem de 20%-30%, sendo que um exame único pode năo evidenciar lesőes precursoras de câncer cervical em até 50% dos casos positivos.23 A citologia quando repetida tem uma sensibilidade máxima de 75%,24,25 e a colposcopia, embora mais sensível para detectar lesőes de alto grau do que a citologia, apresenta um grade erro entre falso-positivos que pode alcançar a marca estratosférica de 55%.26 Um alerta: no milęnio do genoma, para onde vamos? As pesquisas genômicas em todo o mundo estăo mostrando as identidades genéticas de vários microorganismos patogęnicos ao homem (http://www.ncbi.nlm.nih.gov/Genomes/index.html; www.tigr.org). Uma aplicaçăo óbvia da pesquisa genômica é apontar alvos, ou seqüęncias de DNA, para diagnóstico molecular. Na verdade, uma enorme quantidade de trabalhos a respeito de diagnóstico molecular tęm sido produzidos na última década (Figura 2). Embora, alguns avanços práticos tenham sido obtidos, pode-se esperar ainda muitas aplicaçőes que venham a melhorar a eficięncia do diagnóstico em DST. O diagnóstico molecular é baseado no uso de sondas de DNA que tenham total identidade ou grande similaridade de seqüęncia a uma pequena e conservada parte do genoma do organismo que se pretende detectar, geralmente um gene ou uma pequena seqüęncia de um gene deste organismo. Para tanto, geralmente busca-se uma seqüęncia altamente conservada no organismo alvo, os chamados house- keeping genes (HKG) ou ainda seqüęncias de RNA ribossômico. Os HKG săo genes que fazem parte dos eventos metabólicos básicos, como enzimas da via glicolítica clássica, ou genes que codificam proteínas estruturais, como a actina formadora do citoesqueleto. Desta forma, em qualquer situaçăo metabólica, ou em todos os isolados clínicos de qualquer parte do mundo, espera-se que estes genes estejam representados no genoma do microorganismo em questăo. As seqüęncias de RNA ribossômico apresentam ainda a vantagem de estarem representadas em repetidas cópias no genoma, o que geralmente aumenta a sensibilidade analítica da detecçăo, podendo detectar um pequeno número do organismo alvo. Por serem altamente conservados, é necessário avaliar a especificidade clínica, ou seja a capacidade da sonda em năo apenas detectar o patógeno mesmo que em pequena quantidade (o que é útil em amostras escassas), mas também discriminá-lo de outros possíveis microorganismos de similaridade filogenética ou que simplesmente compartilhem o mesmo ambiente. Algumas derivaçőes de técnicas especiais de biologia molecular tęm sido adaptadas para o uso em diagnóstico (Tabela 1). Tabela 1.
Essencialmente, todas elas tęm como base a hibridizaçăo do DNA e a amplificaçăo do
DNA pela técnica da Reaçăo da Polimerase em Cadeia ou PCR. A PCR é uma técnica
poderosa que permite a amplificaçăo exponencial de um pequeno trecho de DNA
limitado pelo par de sondas que o usuário utiliza. Assim, obtęm-se grandes
quantidades de DNA amplificado, sendo possível detectar a presença do DNA de um
organismo mesmo que a amostra seja escassa ou a parasitemia seja ínfima. Essa é a
grande vantagem da PCR sobre a hibridizaçăo de DNA. No entanto, a PCR convencional
necessita de análise em eletroforese com equipamentos e corantes especiais e tóxicos.
Também, a sua interpretaçăo para o diagnóstico baseado na presença ou ausęncia de
DNA amplificado pode ser subjetiva. Ainda, dada a sua enorme sensibilidade, cuidados
adicionais quanto a contaminaçăo entre amostras deve ser levado em conta, até
mesmo a separaçăo física do ambiente, do equipamento e dos manipuladores.
A conjugaçăo da PCR e hibridizaçăo levou a elaboraçăo de técnicas mais refinadas,
com maior sensibilidade, através do uso de uma sonda interna ao DNA amplificado
mais específica que permite maior objetividade na interpretaçăo do resultado e até a
tipagem do organismo alvo. A hibridizaçăo tem evoluído no sentido da marcaçăo das
sondas com compostos năo-radioativos que para as técnicas derivadas, como a PCR-
Dot blot, PCR-ELISA e os chips de DNA, năo comprometem a sensibilidade
analítica. Em termos de DST, a possibilidade de tipagem do organismo alvo fez a PCR-
Dot blot e PCR-ELISA serem implementadas para diagnóstico de tipagem do
HPV.24,27,28 A PCR-ELISA tem vantagem por sua maior automaçăo, e
também tem demonstrado sensibilidade superior que a hibridizaçăo em Dot
blot.27 O fato de năo utilizar eletroforese e adaptar a detecçăo do
produto amplificado em um leitor de ELISA é um atrativo a mais, também sendo
possível a tipagem do organismo em escala e rapidez considerável. Os chips
de DNA29 săo novidades tecnológicas com o mesmo princípio de
hibridizaçăo, porém em pequeno volume e em grande escala, com capacidade analítica
para mais de 100 mil reaçőes simultaneamente. Seu uso restringe-se ŕ pesquisa
genômica, assim como o sequenciamento de DNA.
A PCR-Multiplex utiliza mais de um par de sondas em uma única reaçăo. Teoricamente,
năo há limites para o número de sondas a se utilizar em uma reaçăo de PCR, suprindo-
se dos substratos. Este fato é extremamente interessante quando pensamos em
detectar infecçőes simultâneas, ou microrganismos de relaçăo etiopatogęnica que
compartilhem o mesmo hábitat. Em termos de DST, alguns sistemas interessantes de
PCR-Multiplex tęm sido desenvolvidos para detecçăo simultânea de: Gardnerella
vaginalis e T. Vaginalis;30 T. pallidum, H. ducreyi
e HSV;31 C. trachomatis e N.
gonorrhoeae.32-34 Como muitas DST tęm hábitat delimitado ou
predominante no ambiente cervicovaginal, o desenvolvimento de sondas para PCR-
Multiplex tęm um grande apelo para o elaboraçăo de kits simples de diagnóstico
múltiplo, por isso mais rápidos e de menor custo. Ainda mais, com a maior facilidade
técnica e praticidade da PCR-ELISA, seria um grande avanço o desenvolvimento de um
sistema robusto de PCR-Multiplex conjugado a PCR-ELISA para o diagnóstico
simultâneo de DST e tipagem do HPV.
Năo há dúvidas de que o diagnóstico molecular aumenta substancialmente a eficięncia
da detecçăo da infecçőes genitais, seja qual for a natureza da infecçăo e a técnica de
diagnóstico padrăo implementada.16-18,20,35-43 Năo obstante, as
técnicas convencionais de citologia, microbiologia e imunologia, ainda que
insatisfatórias, serăo sempre o método de escolha até que haja a elaboraçăo de
metodologias que, além de serem mais sensíveis e específicas, também demonstrem
ser rápidas, de fácil execuçăo, de grande processividade e de baixo custo. Neste
contexto, uma das linhas de pesquisa de nosso grupo é o desenvolvimento de novas
tecnologias de diagnóstico para DST, o qual procuramos parceiros da iniciativa privada
que tenham interesse em investir recursos para desenvolvimento de tecnologia
nacional na elaboraçăo de um kit diagnóstico.44
As novas tecnologias de diagnóstico baseadas na detecçăo do DNA do patógeno tęm
evoluído neste sentido. O exemplo mais claro disso resultou em um produto de
diagnóstico de HPV comercialmente disponível que tem atualmente grande uso, a
captura híbrida (Digene Diagnostics, Inc.). Através de um conjunto de sondas para os
diferentes tipos de HPV, a captura híbrida permite a detecçăo semiquantitativa da
carga viral e tipagem entre HPV de baixo risco (tipos 6, 11, 42, 43 e 44) e de alto risco
oncogęnico (tipos 16, 18, 31, 33, 35, 45, 51, 52 e 56). Interessante do processo é a
sua automaçăo, facilitando o laboratorista.
O diagnóstico molecular do HPV traz ŕ superfície um dilema: como, e a que nível
aplicar esta tecnologia na rotina clínica. Trabalhos prospectivos que levam em
consideraçăo a idade da paciente, a chance de infecçăo por HPV, a necessidade e os
custos de exames e tratamentos adicionais, a perda produtiva e estimativa de vida
mostram que haveria reduçăo de custos da ordem de 20%-30% utilizando a captura
híbrida como forma de triagem.3 Esses resultados podem refletir uma
tendenciosidade econômica em prol do consumo da tecnologia de captura híbrida, por
outro lado impőem uma limitaçăo prática ao uso da captura híbrida como rotina de
diagnóstico, em especial para países em desenvolvimento: o custo.
A empresa norte-americana Digene detém a patente da captura híbrida. O
equipamento, tipo fluorômetro, é desenhado para uso exclusivo da captura, năo tendo
outro proveito laboratorial. Os kits de diagnóstico săo caros, embutidos no valor os
royalties e as taxas de importaçăo. A realidade dos países em
desenvolvimento, somado aí as questőes políticas e socio-econômicas, năo p híbrida
em larga escala no sistema público de saúde. Entăo, o diagnóstico pela captura híbrida
fica restrito aos convęnios privados de saúde.
Em tempos em que a Cięncia avança a passos largos, e o Brasil incluído como um dos
grandes líderes mundiais nas Cięncias do Genoma, o que podemos esperar desse
conhecimento em termos de aplicaçăo e benefício sociais, uma vez que o país sofre
com grandes desigualdades socio-econômicas? É certo que năo falta competęncia e
conhecimento científico por parte dos pesquisadores pertencentes ao países em
desenvolvimento. A questăo passa por políticas públicas que priorizem o investimento
em pesquisa nacional. Afinal, com incentivo do governo, temos capacidade de
desenvolver nossos medicamentos e tecnologias de diagnóstico. Estamos certos de que
o custo social será inferior ao que o país desembosa para o pagamento de
royalties, importaçăo de medicamentos e tecnologias de diagnóstico.
2. Over M, and Piot P. HIV infection and sexually transmitted diseases. In: Jamison, D.T., Mosley, W.H., Measham, A.R., Bobadilla, J.L. (Eds.), Disease Control Priorities in Developing Countries 1996; Oxford University Press, New York, USA. 3. Dôres GB, Taromaru EK, Gallo C. Aspectos atuais do rastreamento das lesőes HPV induzidas e do câncer do colo uterino com métodos morfológicos e biomoleculares. Newslab, 1999; 35:196-205. 4. Parker R, and Camargo Jr KR. Poverty and HIV/AIDS: anthropological and sociological aspects. Cad Saúde Pública 2000; 16(suppl.1): 89-102. 5. Brabin L. Clinical management and prevention of sexually transmitted diseases: a review focusing on women. Acta Tropica 2000; 75:53-70. 6. Wasserheit JN. Epidemiological synergy: interrelationships between human imunodeficiency virus infection and other sexually transmitted diseases. STD 1992; 19:61-77. 7. Laga M, Manoka A y col. Non-ulcerative sexually transmitted disease treatment as risk factors for HIV-1 transmission in women: results from a cohort study. AIDS 1993; 7:95-102. 8. Sewankambo N, Gray RH y col. HIV-1 infection associated with abnormal vaginal flora morphology and bacterial vaginosis. The Lancet 1997; 350: 546-550. 9. Dallabeta G, and Diomi C. Treating sexually transmitted diseases to control HIV transmission. Curr Opin Infect Dis 1997; 10:22-25. 10. Simőes-Barbosa1 A., Feijó GC, Silva JX, Barbosa TW. A Six-years Follow up Survey of Sexually Transmitted Diseases in Brasilia, Capital of Brazil. Braz J Inf Dis 2002; 6(3): 110-117. 11. WHO. Bulletin of WHO. 1996; 74(4): 345-351. 12. WHO. Cervical Cancer Screening: Managerial Guidelines 13. Amaral, E. Current approach to STD management in women. Int J Gynecol Obst 1998; 63(Suppl.1):S183- S189. 14. Gompel C, and Koss LG. Citologia ginecológica e suas bases anatomoclínicas. 1997; Editora Manole Ltda, Săo Paulo, Brasil. 15. Koss LG. The Papanicolaou test for cervical cancer detection: a triumph and a tragedy. JAMA 1989; 261:737-743. 16. Lobo1 TT, Feijó G y col. A comparative evaluation of the Papanicolaou for diagnosis of Trichomonosis. Sex Transm Dis 2003; 30(9): 694-699. 17. Lobo2 TT, Feijó G, and Simőes-Barbosa A.. Relato de um estudo comparativo de diagnóstico da tricomonose no Distrito Federal. Newslab 2002; 53: 156-162. 18. Tong CY, Mallinson H. Moving to nucleic acd-based detection of genital Chlamydia trachomatis. Expert Rev Mol Diagn 2002; 2:257-266. 19. Larsen SA., Steiner BM, and Rudolph AH. Laboratory diagnosis and interpretation of tests for syphilis. Clin Microbiol Rev 1995; 8-1-21. 20. Bowden FJ, Tabrizi SN, Garland SM, Fairley CK. Infectious diseases. 6: Sexually transmitted infectious: new dianostic approaches and treatments. Med J Aust 2002; 176:551-557. 21. Hunter H, and Handsfield MD. Color Atlas and Synopsis of Sexually Transmitted Diseases 2000; 216 pp. McGraw-Hill Professional Publishing. New York, N.Y. 22. Sherman ME, Schiffman MH y col. Toward objective quality assurane in cervical cytopathology: correlation of cytopathologic diagnoses with detection of high-risk human papillomavirus types. Am J Clin Pathol 1994; 102:182-187. 23. Ferenczy A. HPV, colposcopy and cytology? HPV summit 1999; pag. 3. 24. Mannos MM, Kinney WK y col. Identifying women with cervical neoplasia: using human papillomavirus DNA testing for equivocal Papanicolaou results. JAMA 1999; 281:1605-1610. 25. Wright TC, Sun XW, Koulos J. Comparison of management algorithms for the evaluation of women with low-grade cytologic abnormalities. Obstrect Gynecol 1995; 85: 202-210. 26. Mitchell MF, Schottenfeld D y col. Colposcopy for the diagnosis of squamous intraepithelial lesions: a meta- analysis. Obstet Gynecol 1998; 91:626-631. 27. Poljak M, Seme K. Rapid detection and typing of human papillomaviruses by consensus polymerase chain reaction and enzyme-linked immunosorbent assay. J Virol Methods 1996; 56:231-238. 28. Mannos MM, Ting Y y col. Use of polymerase chain reaction amplification for the detection of genital human papillomaviruses. Cancer Cells 1989; 7:209-214. 29. Brown TA. Sequencing genomes. In: Brown, T.A. Genomes. John Wiley & Sons Inc., New York, NY 1999; 59-84. 30. Briselden AM, and Hillier SL. Evaluation of Affirm VP Microbial Identification Test for Gardnerella vaginalis and Trichomonas vaginalis. J Clin Microbiol 1994; 32:148-152. 31. Orle KA, Gates CA y col. Simultaneous PCR detection of Haemophilus ducreyi, Treponema pallidum, and herpes simplex virus types 1 and 2 from genital ulcers. J Clin Microbiol 1996; 34:49-54. 32. Farrel DJ. Evaluation of AMPLICOR Neisseria gonorrhoeae PCR uisng cppB nested PCR and 16S rRNA PCR. J Clin Microbiol 1999; 37:386-390. 33. Van der Pol B. COBAS Amplicor: na automated PCR system for detection of Chlamydia trachomatis and Neisseria gonorrhoeae. Expert Rev Mol Diagn 2002; 2:379-389. 34. Leslie DE, Azzato F, Ryan N, Fyfe J. An asessment of the Roche Amplicor Chlamydia trachomatis/Neisseria gonorrhoeae multiplex PCR assay in routine diagnostic use on a variety of specimen types. Commun Dis Intell 2003; 27:373- 379. 35. Buimer M, Van Doornum GJJ, y col. Detection of Chlamydia trachomatis and Neisseria gonorrhoea by ligase chain reaction-based assays wiht clinical specimens form various sites: implications for diagnostic testing and screening. J Clin Microbiol 1996; 34:2395-2400. 36. Caul EO, Horner PJ, y col. Population-based screening programmes for Chlamidia trachomatis. Lancet 1997; 349:1070-1071. 37. Fairley CK, Bowden FJ, y col. Sexually transmitted diseases in disadvantaged Australian communities. JAMA 1997; 278:117-118. 38. Stary A. Chlamydia screening: which sample for which tecnhique? Genitourin Med 1997; 73: 99-102. 39. Madico G, Quinn TC y col. Diagnosis of Trichomonas vaginalis infection by PCR using vaginal swab samples. J Clin Microbiol 1998; 36(11):3205-3210. 40. Van der Schee C, Van Belkun A y col. Improved diagnosis of Trichomonas vaginalis infection by PCR using vaginal swabs and urine specimens compared to diagnosis by wet mount microscopy, culture, and fluorescent staining. J Clin Microbiol 1999; 37(12):2127-4130. 41. Mayta H, Gilman RH y col. 18S Ribosomal DNA-based PCR for diagnosis of Trichomonas vaginalis. J Clin Microbiol 2000; 38(7):2683-2687. 42. Press N, Chavez VM y col. Screening for sexually transmitted diseases in Human Immunodeficiency Virus- positive patients in Peru reveals an absence of Chlamydia trachomatis and identifies Trichomonas vaginalis in pharyngeal specimens. Clin Infec Dis 2001; 32:808-814. 43. Lewis DA. Chancroid: clinical manifestations, diagnosis, and management. Sex Transmi Infect 2003; 79:68-71. 44. Simőes-Barbosa2 A, Lobo T, Feijó GC. Em busca de Parcerias para um Novo Produto de Diagnóstico de
|
|
|
|
|
 |
Columnista
Experto de SIIC Tanri Shiozawa Profesor asociado, Departamento de Ginecología y Obstetricia, Escuela de Medicina de la Universidad Shinshu, Asahi, Japón.*En colaboración con los doctores Toshio Nikaido e Ikuo Konishi. |
|
|
|
Trabajos Distinguidos, Obstetricia y Ginecología , integra el Programa SIIC de Educación Médica Continuada |
![]()
Bienvenidos
a siicsalud
Acerca de SIIC Estructura de SIIC
Sociedad Iberoamericana de Información
Científica (SIIC)
Av. Belgrano 430, (C1092AAR), Buenos Aires, Argentina
Tel: +54 11 4342-4901; Fax: +54 11 4331-3305.
atencionallector@siicsalud.com
Casilla de Correo 2568, (C1000WAZ) Correo Central, Buenos
Aires.
Copyright siicsalud© 1997-2004, Sociedad Iberoamericana de Información Científica (SIIC)