

-
 Alergia
Alergia -
 Anestesiología
Anestesiología -
 Bioquímica
Bioquímica
-
 Cardiología
Cardiología
-
 Cirugía
Cirugía
-
 Dermatología
Dermatología
-
 Endocrinología y Metabolismo
Endocrinología y Metabolismo
-
 Enfermería
Enfermería
-
 Gastroenterología
Gastroenterología
-
 Hematología
Hematología
-
 Infectología
Infectología
-
 Inmunología
Inmunología
-
 Medicina Interna
Medicina Interna
-
 Nefrología
Nefrología
-
 Neumonología
Neumonología
-
 Neurología
Neurología
-
 Nutrición
Nutrición
-
 Obstetricia y Ginecología
Obstetricia y Ginecología
-
 Odontología
Odontología
-
 Oncología
Oncología
-
 Otorrinolaringología
Otorrinolaringología
-
 Pediatría
Pediatría
-
 Salud Mental
Salud Mental
-
 Salud Pública
Salud Pública
-
 Urología
Urología
-
 Más Especialidades
Más Especialidades
- Índice
-
Acceso por especialidad
-
Alergia
-
Anestesiología
-
Bioquímica
-
Cardiología
-
Cirugía
-
Dermatología
-
Endocrinología y Metabolismo
-
Enfermería
-
Gastroenterología
-
Hematología
-
Infectología
-
Inmunología
-
Medicina Interna
-
Nefrología
-
Neumonología
-
Neurología
-
Nutrición
-
Obstetricia y Ginecología
-
Odontología
-
Oncología
-
Otorrinolaringología
-
Pediatría
-
Salud Mental
-
Salud Pública
-
Urología
-
Más Especialidades
-
Noticias biomédicas
-
Fuentes informativas
- Editoriales
-
COVID-19
-
INFORMES CIENTÍFICOS
- Preguntas con Respuesta
- América Latina Investiga
- Completos revisados (full text)
-
NOTICIAS/OPINIONES
-
Expertos invitados
- Por especialidad
- de Iberoamérica
- del mundo
- Entrevistas
- Casos clínicos
-
Crónicas de Autores
- Authors’ Reports
-
Instrucciones para autores
-
Expertos Preguntan
-
Novedades
- ViASIIC
- Medicina del Dolor
-
Por especialidad
-
Iberomédica
-
Informes comentados
- Textos Completos Autorizados
- Preocupaciones Profesionales
-
Red Científica Iberoamericana (RedCIbe)
-
Conceptos Categoricos
-
Acceso por especialidad

-
Alergia
-
Anestesiología
-
Bioquímica
-
Cardiología
-
Cirugía
-
Dermatología
-
Endocrinología y Metabolismo
-
Enfermería
-
Gastroenterología
-
Hematología
-
Infectología
-
Inmunología
-
Medicina Interna
-
Nefrología
-
Neumonología
-
Neurología
-
Nutrición
-
Obstetricia y Ginecología
-
Odontología
-
Oncología
-
Otorrinolaringología
-
Pediatría
-
Salud Mental
-
Salud Pública
-
Urología
-
Más Especialidades
-
Noticias biomédicas
-
Fuentes informativas
- Editoriales
- Covid-19
-
Informes Científicos
- Preguntas con Respuesta
- América Latina Investiga
- Completos revisados (full text)
- Noticias/Opiniones
- Noticias (castellano/portugués)
- Noticias (otros idiomas)
- Expertos invitados
- Por especialidad
- de Iberoamérica
- del mundo
- Entrevistas
- Casos clínicos
-
Crónicas de Autores
- Authors’ Reports
-
Instrucciones para autores
-
Expertos Preguntan
- Novedades
- ViASIIC
- Medicina del Dolor
-
Por especialidad
-
Iberomédica
-
Informes comentados
- Textos Completos Autorizados
- Preocupaciones Profesionales
- Red Científica Iberoamericana
- Por materia
- Por fecha
-
Conceptos Categóricos

(especial para SIIC © Derechos reservados)
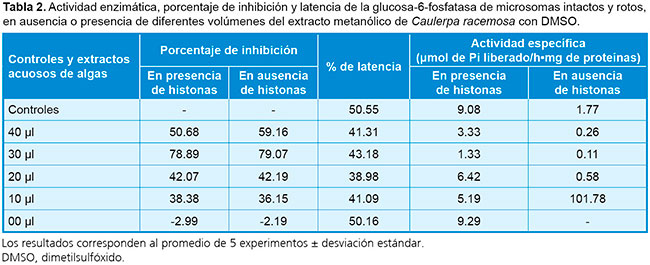
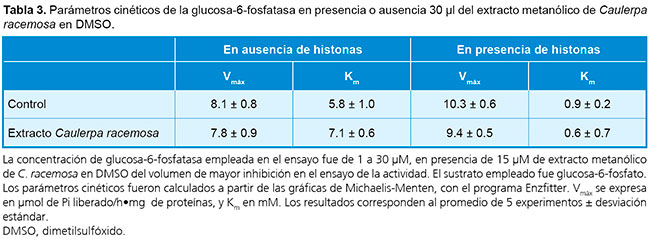
Por las distintas complicaciones que reviste a nivel mundial la diabetes mellitus tipo 2 en los pacientes, resulta de interés la búsqueda de nuevas alternativas terapéuticas. Este estudio encontró un efecto inhibitorio significativo sobre el transportador T1 y la subunidad catalítica, que constituyen parte importante del sistema glucosa-6-fosfatasa, por el extracto metanólico del alga Caulerparacemosa, lo que orienta a la presencia de metabolitos polares capaces de ejercer una posible acción antihiperglucemiante.
Daniel Enrique Ríos Acosta
Columnista Experto de SIIC
Institución:
Universidad Católica del Maule Artículos publicados por Daniel Enrique Ríos Acosta
Freddy González Mujica* Juan Linares**
Médico cirujano, Instituto de Medicina Experimental, Caracas, Venezuela*
Biólogo, Universidad Pedagógica Experimental Libertador, Caracas, Venezuela**
|
|
|
Introducción: La diabetes mellitus tipo 2 es una de las enfermedades más frecuentes y con más complicaciones metabólicas que está presente en nuestros días. Se caracteriza por hiperglucemia persistente que puede conducir a daños en órganos, por lo que su realidad multisistémica representa un reto global en salud. La enzima glucosa-6-fosfatasa cataliza la última etapa de la neoglucogénesis y la glucogenólisis, formando parte del mecanismo homeostático de la glucosa en el torrente sanguíneo. Objetivo: Estudiar el efecto del extracto de algas sobre la actividad de la glucosa-6-fosfatasa existente en microsomas de hígado de rata de la cepa Sprague-Dawley. Métodos: Se realizó una investigación exploratoria de campo para analizar las propiedades antihiperglucemiantes del extracto de algas Caulerpa racemosa, Laurencia sp., Pterocladiella capilacea y Grateloupia doriphora. Estas especies fueron colectadas en la playa del estado La Guaira, Venezuela, durante el período enero-abril de 2020. A partir de los extractos obtenidos, se evaluó la actividad inhibitoria sobre la enzima glucosa-6-fosfatasa existente en microsomas hepáticos de ratas. Resultados: Entre los extractos analizados, se destacó el metanólico en DMSO de Caulerpa racemosa (especie invasora en Venezuela), con un porcentaje de inhibición en presencia de proteínas histónicas de 78.89%, y en su ausencia, un porcentaje de 79.07%. Conclusiones: Estos hallazgos sugieren la existencia de metabolitos secundarios polares en esta alga, capaces de reaccionar con mayor acción sobre la subunidad catalítica de la glucosa-6-fosfatasa inhibiendo la capacidad neoglucogénica de los microsomas. antihiperglucemiante, Caulerparacemosa, diabetes mellitus tipo 2, glucosa-6-fosfatasa, microsomas
Artículo completo
EFECTO DE EXTRACTOS ALGALES SOBRE LA ACTIVIDAD DE LA GLUCOSA-6-FOSFATASA PRESENTE EN MICROSOMAS DE HIGADO DE RATAS
1. Harreiter J, Roden M. Diabetes mellitus: definition, classification, diagnosis, screening and prevention. WienKlinWochenschr 135(suppl1):7-17, 2023. 2. ElSayed NA, Aleppo G, Aroda VR, Bannuru RR, Brown FM, et al. Classification and diagnosis of diabetes: standards of care in diabetes-2023. Diabetes Care 46(suppl 1):19-40, 2023. 3. Chiquete J, Nuño P, Panduro A. Perspectiva histórica de la diabetes mellitus: comprendiendo la enfermedad. InvestigaciónenSalud 3:5-10, 2001. 4. Widnell C, Tata J. A procedure for the isolation of enzymically active rat-liver nuclei.Biochem J 92(2):313-317, 1964. 5. González-Mujica F. Glucosa-6-fosfatasa de envoltura nuclear de hígado de ratas. InvestClínica 49(2):169-180, 2008. 6. Foster J, Nordlie R. The biochemistry and molecular biology of the glucose-6-phosphatase system.E Biol Med 227(8)601-608, 2002. 7. Bocanegra A, Macho González A, Garcimartín A, Bendi J, Sánchez-Muniz F. Whole alga, algal extracts, and compounds as ingredients of functional foods: composition and action mechanism relationships in the prevention and treatment of type-2 diabetes mellitus. Int J MolSci 22(8):3816, 2021. 8. Sun Z, Chen F. Evaluation of the green alga Chlorella pyrenoidosa for management of diabetes. J Food Drug Anal 20(suppl 1):246-249, 2012. 9. Lauritano C, Ianora A. Marine organisms with anti-diabetes properties. Mar Drugs 14(12):220, 2016. 10. Qudus B, Aroyehun A, Abdul Razak S, Palaniveloo K, Nagappan T, et al. Bioprospecting cultivated tropical green algae, Caulerparacemosa (Forsskal) J. Agardh. A perspective on nutritional properties, antioxidative capacity and anti-diabetic potential. Foods 9(9):1313, 2020. 11. Dissanayake IH, Bandaranayake U, Keerthirathna LR, Manawadu C, Silva RM, et al. Integration of in vitro and in-silico analysis of Caulerparacemosa against antioxidant, antidiabetic, and anticancer activities. Sci Rep 12(1):20848, 2022. 12. Permatasari HK, Nurkolis F, Hardinsyah H, Taslim NA, Sabrina N, et al. Metabolomic assay, computational screening, and pharmacological evaluation of Caulerparacemosa as an anti-obesity with anti-aging by altering lipid profile and peroxisome proliferator-activated receptor-B3; coactivator 1-B1; Levels. Front Nutr 9:939073, 2022. 13. Marcucci O, González-Mujica F, Perez-Ayuso E. Alterations of liver nuclear envelopes accompanying thioacetamide administration in rats. Acta CientVenez 34(2):109-117, 1983. 14. González-Mujica F, Motta N. Actividad antihiperglicemiante de Bauhiniamegalandra. Vitae 43, 2010. 15. Burchell A, Hume R, Burchell B. A new microtechnique for the analysis of the human hepatic microsomal glucose-6-phosphatase system.ClinChimActa 173(2):183-192, 1988 16. Taussky HH, Shorr E, Kurzmann G. A microcolorimetric method for the determination of inorganic phosphorus. J BiolChem 202(11):675-685, 1953. 17. Arion WJ, Schulz LO, Lange AJ, Telford JN, Walls HE. The characteristics of liver glucose-6-phosphatase in the envelope of isolated nuclei and microsomes are identical. J BiolChem 258(9):12661-12669, 1983. 18. Igarashi Y, Kato S, Narisawa K, Tada K, Amano Y, et al. A direct evidence for defect in glucose-6-phosphate transport system in hepatic microsomal membrane of glycogen storage disease type IB. BiochemBiophys Res Commun 119(2):593-597, 1984. 19. Westergaard N, Madsen P, Lundbeck JM, Jakobsen P, Varming A, Andersen B. Identification of two novel and potent competitive inhibitors of the glucose-6-phosphatase catalytic protein. Diabetes ObesMetab 4(2):96-105, 2002. 20. Singh J, Nordlie RC, Jorgenson RA. Vanadate: a potent inhibitor of multifunctional glucose-6-phosphatase. BiochimBiophys Acta 678(3):477-482, 1981. |
|


|
Está expresamente prohibida la redistribución y la redifusión de todo o parte de los contenidos de la Sociedad Iberoamericana de Información Científica (SIIC) S.A. sin previo y expreso consentimiento de SIIC. |

